Article Text

Abstract
Exosomes, as the main group of extracellular vesicles, are biologically active lipid-bilayer vesicles that are naturally released from different types of normal or tumor cells. These vesicles play an important role in intercellular communication and influence the extracellular environment and the immune system. Emerging evidence demonstrates that cancer-derived exosomes are enriched in immunosuppressive proteins, such as the programmed death-ligand 1 (PD-L1). PD-L1 and its receptor programmed cell death protein 1 (PD-1) are the key immune checkpoint molecules that promote tumor progression via negative regulation of immune responses. PDL-1 is highly expressed on the surface of tumor cells and binds to PD-1 on the surface of activated T cells, leading to suppression of T cells, which consequently enables cancer cells to escape antitumor immunity. Currently, there are several Food and Drug Administration-approved monoclonal antibodies blocking PD-1/PD-L1 interaction, which are clinically used for cancer treatment. However, despite impressive treatment outcomes, some patients show poor response to PD-1/PD-L1 blockade. Of note, tumor-derived exosomes containing PD-L1 can recapitulate the effect of cell-surface PD-L1. There is evidence that reveals a significant association between levels of circulating exosomal PD-L1 and rate of response to anti-PD-1/PD-L1 antibody therapy. The present article reviews the role of exosomal PDL-1 in the therapeutic resistance to anti-PD-1/PD-L1 treatment. Importantly, it is suggested that the removal of exosomal PDL-1 could serve as a therapeutic adjuvant for enhancing the efficacy of anti-PD-1/PD-L1 therapy in patients with cancer.
- programmed cell death 1 receptor
- immunotherapy
- tumor escape
This is an open access article distributed in accordance with the Creative Commons Attribution Non Commercial (CC BY-NC 4.0) license, which permits others to distribute, remix, adapt, build upon this work non-commercially, and license their derivative works on different terms, provided the original work is properly cited, appropriate credit is given, any changes made indicated, and the use is non-commercial. See http://creativecommons.org/licenses/by-nc/4.0/.
Statistics from Altmetric.com
PD-1/PD-L1 immune checkpoint pathway as a double-edged sword
Immune checkpoints are cell surface regulatory receptors, located mainly, but not exclusively, on T lymphocytes. After recognizing cognate ligands on the antigen-presenting cells or the target cells, these receptors act as T cell receptor (TCR) cosignaling partners that deliver either stimulatory or inhibitory signals to regulate the lymphocyte activation.1
Programmed cell death protein 1 (PD-1) is a key immune checkpoint molecule known as one of the major inhibitory coreceptors expressed in the activated T cells.2 Under normal physiology, immune checkpoints, such as PD-1, are critical for maintaining self-tolerance, preventing autoimmunity, and controlling T-cell responses within a desired physiological range to protect tissues from excessive inflammatory reactions. However, in cancerous conditions, these regulatory proteins can allow tumor cells to protect themselves from the antitumor T-cell responses, causing the so-called tumor immune evasion.1
PD-1 acts through interaction with PD-L1 that is a highly expressed ligand in non-lymphoid tissues in response to inflammatory cytokines such as interferon-gamma (IFN-γ) produced by cytotoxic T Cells.2 3 On engagement with PD-L1, PD-1 transmits a negative costimulatory signal in T cells through the recruitment of Src homology 2 domain containing phosphatases 1/2 (SHP1/2) that dephosphorylates downstream TCR-mediated signaling elements and thereby inhibits T cell proliferation, cytokine production and release, and cytotoxicity.4 Therefore, the PD-1/PD-L1 regulatory system is induced by immune responses and then through a negative feedback loop attenuates T-cell responses and minimizes tissue damage.
On tumor antigen recognition by T cells, the released IFN-γ triggers tumor cells to express PD-L1 that binds coinhibitory receptor molecule PD-1 on cytotoxic T cells within the tumor microenvironment (TME). This allows the specific inhibition of tumor recognition by T cells, resulting in attenuating antitumor immune responses and promoting tumor growth. IFN-γ induces the expression of PD-L1 through activating signaling pathways downstream of the type II interferon receptor, including Janus kinase and signal transducer and activators of transcription, leading to binding of the IRF1 transcription factor to the PD-L1 gene promoter.5 Moreover, oncogenic pathways, including MAPK (RAS/RAF/MEK/ERK) and PI3K/Akt/mTOR, are also known to mediate the induction of PD-L1 expression through activating c-JUN that, as the component of AP-1 transcription factor, binds to the enhancer element on PD-L1 gene and augments the transcription signal in tumor cells.6
Therapeutic effects of PD-1/PD-L1 blocking antibodies
Immune checkpoint blockade therapies are now approved by US Food and Drug Administration (US-FDA) for the treatment of a wide range of cancer types, with approval likely for additional indications in the near future. Among negative immune checkpoint molecules, the blockade of PD-1 and its major ligand, PD-L1, provides one of the most successful immunotherapies by intensifying T cell immunity against cancer cells. Since the 2011 FDA approval of the first immune checkpoint inhibitor, ipilimumab (anti-CTLA4), for the treatment of metastatic melanoma, six monoclonal antibodies targeting the PD-1 (pembrolizumab, nivolumab, and cemiplimab) and PD-L1 (atezolizumab, durvalumab, and avelumab), have been approved for the treatment of various cancer types (table 1).1
FDA-approved monoclonal antibodies blocking immune checkpoints in human cancer
An important rationale respecting the development of anti-PD-1/PD-L1 drugs for cancer treatment emerged from the key findings that showed PD-1 is over-expressed on tumor-infiltrating lymphocytes (TILs),7 8 while PD-1 ligand is highly upregulated in most of the human cancer cells.9 This was validated through many preclinical studies that indicated mAbs blockade of PD-1 or PD-L1 significantly improved antitumor immunity.9–11 Anti-PD-1/PD-L1 blocking mAbs have heightened tumor selectivity and decreased toxicity as well as a much broader spectrum of antitumor activity when compared with anti-CTLA-4 mAbs. There is evidence that clinical responses to immune checkpoint inhibitors might be correlated with immune-related adverse events (irAEs). Overall, drug-induced irAEs are more likely to be experienced in patients treated with anti-CTLA-4 (60%–85%) than anti-PD-1 (16%–37%) or anti-PD-L1 (12%–24%).12–14
Pembrolizumab, the first mAbs against PD-1, achieved its first global approval for patients with unresectable or metastatic melanoma by US-FDA.15 Afterward, its use was extended to non-small cell lung cancer (NSCLC),16 head and neck squamous cell carcinoma (HNSCC),17 cervical carcinoma,18 and metastatic urothelial carcinoma18 among others in a list that continues to grow. Anti-PD-1 antibodies can block the PD-1 signaling pathway, thus preventing PD-1-mediated attenuation of TCR signaling, which promotes reinvigoration of exhausted PD-1+ CD8+ T cells, resulting in immune tumor rejection.19–22 Although the precise molecular and cellular events mediating enhancement of antitumor immunity by PD-1 blockade are not fully understood, tumor neoantigen-specific CD8+ T cells appear to be the major T-cell population mediating anti-PD-1 responses.23 Apart from restoring T-cell activity through modulation of TCR signaling, blockade of the PD-1 signaling axis is also able to reverse the associated metabolic reprogramming to an extent which in part mediates the reinvigoration of tumor antigen-specific T cells.24 25
Alongside direct inhibition of PD-1, antibodies targeting PD-L1 are also sufficient to reverse the negative immune regulation and, thereby, reinvigorate the host antitumor immunity. The current FDA-approved mAbs targeting PD-L1 are known to be efficient for treating several cancer types, including NSCLC, small cell lung cancer, urothelial bladder cancer, breast cancer, and Merkel cell carcinoma.1 In the light of the dominance in PD-L1 expression, blockade of PD-L1 can reiterate the impact of PD-1 blockade. PD-L1 expression is primarily triggered by Th1 cytokines, such as IFN-γ. This can in part describe the effectiveness of PD-L1 blockade since Th1-skewed responses would be more desirable for inducing antitumor immunity.26 By contrast with PD-1 blockade, anti-PD-L1 antibodies may also achieve part of their efficacy from antibody-dependent cellular cytotoxicity. This claim emerged from an in vivo study that shows Fc receptor binding is crucial for the effectiveness of anti-PD-L1, but not anti-PD-1, antibody therapy-promoted tumor regression in mice bearing tumor.27 Notably, these findings suggest that anti-PD-1 and anti-PD-L1 antibodies have distinct biological activities and act, at least in part, through different mechanisms.
Factors modulating response to PD-1/PD-L1 inhibitors
Despite the above-mentioned advancements in clinical practice, some patients and cancer types show therapeutic resistance or relatively low-response rate to PD-1/PD-L1 inhibitors.28–30 The basis of differential therapeutic success between patients and between cancers is not completely understood. However, several factors, such as the TME, the tumor genomics, and systemic factors such as exosomes have been suggested for explaining potential mechanisms of the response and resistance to PD-1/PD-L1 inhibitors.
Of TME-related factors, PD-L1 expression status and pre-existing TILs have been widely studied.20 31 Although the conclusions from various clinical trials are not consistent, a significant positive but not absolute correlation between PD-L1 expression in the TME and responsiveness to anti-PD-1/anti-PD-L1 therapy has been generally found.32 33 Across all tumor types, patients with PD-L1-negative tumors respond to anti-PD-1/PD-L1 therapy in 0%–17%, while those with PD-L1-positive tumors exhibit a response rate ranging from 44% to 100%.34 Besides, TILs are the other cell types affecting the tumor immune microenvironment. The density and phenotype of pre-existing TILs within a TME can impact the efficacy of anti-PD-1/PDL1 therapy.35 The results from a model of tumor immune microenvironment that consists of TILs and PD-L1 expression status indicate that patients with cancer with PD-L1+ TIL+ tumors possess an effective immune response to PD-1/PD-L1 blockade therapy.36 In patients with NSCLC, pretherapy levels of the intratumoral population of CD8+ cytotoxic T cells with the highest PD-1 expression have shown a positive correlation with anti-PD-1 response.37 Moreover, the ratio of memory-like to exhausted TILs, as two major intratumoral CD8+ T cell phenotypes, has been indicated to be strongly correlated with increased survival and response in a cohort of patients with melanoma treated with anti-PD-1.38
Another studied factor is the tumor mutation burden (TMB). The primary targets of many tumor immune responses are tumor-specific neoantigen peptides that arise from somatic mutations in tumor genomes. The number of non-synonymous single nucleotide variants in a tumor, referred to as TMB, can affect the odds of generating immunogenic peptides and thereby influence response to immune checkpoint blockade in patients.25 39–44 Of note, a higher TMB favors positive response to PD-1/PD-L1 blockade in a variety of tumor types including NSCLC,23 45–48 small cell lung cancer,48 urothelial carcinoma,49 metastatic melanoma,50 and HNSCC.51–53 Moreover, data from a meta-analysis across 27 tumor types show a positive correlation between average response rate and TMB.54 These findings make clear that there is a significant pan-cancer association between TMB and response to PD-1/PD-L1 blockade by the tumor type. Further supporting the relationship between TMB and high sensitivity to PD-1/PD-L1 blockade is the observation that microsatellite instability (MSI)-high/mismatch repair deficiency (MMRd) associates with satisfactory treatment effects in patients with multiple cancer receiving anti-PD-1/PD-L1 therapy.55 Tumors with high MSI generate many neopeptides owing to the hypermutated phenotype. MSI-positive tumors are a specific type of high TMB tumors, with MMRd generating a high mutational load. Notably, MMRd derives several insertion and deletion mutations resulting in frameshifts producing neoantigens that may be more immunogenic due to their higher sequence divergence from self-peptides.55–57 Taken together, these findings reveal that MMRd is correlated with enhanced response to PD-1/PD-L1 blockade because of increased TMB.
Besides, systemic factors are also found to influence the therapeutic efficacy of PD-1/PD-L1 inhibitors. Among them, peripheral blood parameters including neutrophil/lymphocyte ratio, total lymphocyte and monocyte count, relative eosinophil count, T cell clonality, PD-L1high circulating tumor cells, and circulating PD-L1 have been reported to be significantly correlated with PD-1/PD-L1 blockade response in multiple studies of anti-PD-1/PD-L1 therapy across a wide range of cancer types.58 Exosomal PD-L1 is another systemic factor that has recently attracted extensive attention. Tumor cells release PDL1-containing exosomes that have been known to be in part responsible for resistance to anti-PD-1/PDL-1 therapy. A growing body of evidence indicated that exosomal PDL-1 affects the TME and antitumor immunity. In the present review article, we seek impacts of exosomal PD-L1 on clinical outcomes with PD-1/PDL-1 blockade therapy and suggest potential combination therapies to overcome the resistance.
Activity of exosomal PD-L1
Exosomes, as the main group of secreted small extracellular vesicles, are biologically active lipid-bilayer vesicles with a size around 30–100 nm that are naturally produced and released by different types of normal and tumor cells.59 60 These vesicles play an important role in intercellular communication and influence the extracellular environment and the immune system responses.59 61 Exosomes are secreted into extracellular space through endosomal pathways and transport various bioactive molecules (cargo) to the target cells. The composition of exosomal cargo is very diverse and includes a wide range of immunosuppressive and immunostimulatory proteins, chemokines, cytokines, cellular receptors, lipids, as well as different nucleic acids such as micro-RNAs and circular RNAs.62–64
Tumor cells can actively produce large levels of exosomes enriched in cancer-promoting cellular contents, such as immunosuppressive proteins like PD-L1, mRNAs, and micro-RNAs, that participate in cancer development and metastasis, through dysregulating antitumor or protumor immunity responses and promoting drug resistance.65–67 Of note, tumor-secreted exosomes contain PD-L1 presented both on the surface and within exosome particles. ESCRTs (endosomal sorting complex required for transport) are involved in the packaging of biomolecules into exosome, and nSMase2 (neutral sphingomyelinase 2) and Rab proteins regulate exosome secretion.68 ESCRT, Rab27a, and nSMase2 have been identified to participate in the packaging and secretion of exosomal PD-L1.69 Exosomes can transport PD-L1 to other cells with low or no PD-L1 expression, with the potential to bind to PD-1.70
The plasma/serum levels of PD-L1 expressed on exosomes, but not soluble PD-L1, are found to associate with disease progression and clinicopathological features in patients with cancer, such as HNSCC71 and NSCLC.72 Of note, major histocompatibility complex (MHC) molecules expressed on exosomes can play an essential role in exosomal PD-L1-mediated tumor promotion. Indeed, molecular interaction of exosomal MHC I with TCR enhances the inhibitory effect of exosomal PD-L1 to T cells. These might be perfectly justifiable reasons for the higher immunosuppressive impact of exosomal PD-L1 than the soluble form.71 72
Exosomal PD-L1 can recapitulate the effect of cell-surface PD-L1; it similarly provokes tumor progression mainly through enabling cancer cells to escape antitumor immunity via inhibiting T cell activation (figure 1). Generally, B cell presentation of antigen to T cells will activate T cells. An in vitro setting of PD-L1 negative Raji B cell and PD-1 expressing Jurkat T cell revealed that exosomal PD-L1 can suppress T cell activation.73 The immunosuppressive effect of tumor-derived exosomal PD-L1 has been widely confirmed by other investigations. Malignant glioma cells have been found to generate exosomes containing PD-L1 that involves in tumor progression. Exosomal PD-L1 secreted by glioblastoma stem-like cells were identified to participate in T cell immunosuppression by inhibition of CD4+ and CD8+ T cell activation.74 PD-L1-containing exosomes isolated from NSCLC patients could inhibit the activity of CD8+ T cells by reducing the production of interleukin 2 (IL-2) and IFN-γ in a dose-dependent manner. Of note, exosomes with high levels of PD-L1 (PD-L1high exosomes) showed a strong potency to suppress the production of IL-2 and IFN-γ, whereas exosomes with lower levels of PD-L1 (PD-L1low exosomes) exerted no significant effect. PD-L1high exosomes could also significantly induce apoptosis in CD8+ T cells through PD-1/PD-L1 interaction.75 Furthermore, in patients with HNSCC, exosomal PD-L1 were able to suppress the proliferation of CD4+ T cells, induce apoptosis in CD8+ T cells, and enhance the suppressor activity of Treg cells, depending on the level of PD-L1 in exosomes. PD-L1high exosomes were found to be more effective in the suppression of T cells, compared with PD-L1low exosomes.71 An in vivo study revealed that exosomal PD-L1 isolated from murine (SCCVII) and human (SCC90) HNSCC cell lines could enhance tumor progression in a mouse model of human oral-oesophageal cancer through reducing migration of CD4+ and CD8+ T cells toward the tumor site.76 Further studies on a mouse model of prostate cancer exhibits that tumor-secreted exosomal PD-L1 can travel to the tumor’s draining lymph nodes where it inhibits T cell activation and eventually leads to T cell exhaustion and reducing spleen size.73 In sum, such experimental and clinical studies strongly emphasis that exosomal PD-L1 through reprinting function of cell-surface PD-L1 can inhibit T cell immunity and enhance tumor growth in different tumor types, including prostate, head and neck, oral-oesophageal, and colorectal cancer.
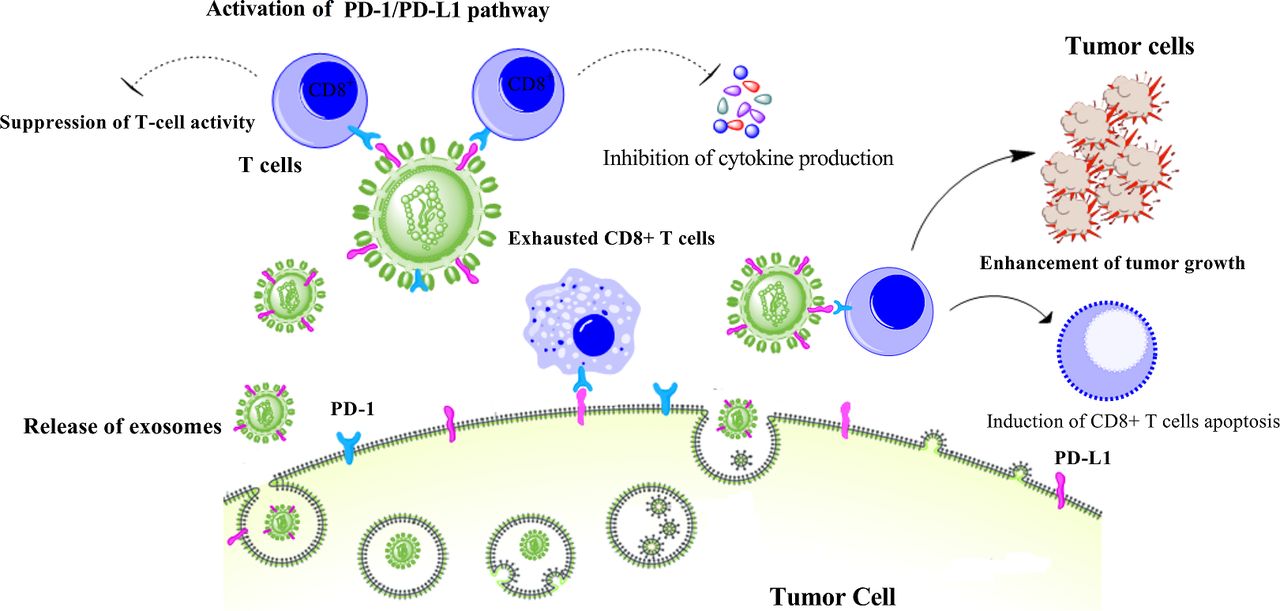
The role of exosomal PD-L1 in cancer progression. Tumor cells secrete exosomes containing PD-L1 leading to suppression of immunity by reducing T-cell activity and inhibition of interferon-gamma and interleukin 2 production as well as reducing total number of CD8+ T cells by inducing apoptosis through PD-1/PD-L1 pathway. PD-1, programmed cell death protein 1; PD-L1, programmed death-ligand 1.
PD-L1 in exosomes and the acquired resistance to anti-PD-1/PD-L1 therapy
There is convincing preclinical and clinical evidence that shows the effective role of exosomal PD-L1 in the relatively low response rate of anti-PD-L1/PD-1 therapy. Clinical significance of plasma circulating exosomal PD-L1 has been indicated in patients with head and neck cancer,71 77 gastric cancer,78 79 NSCLC,72 pancreatic cancer,80 and melanoma.69 Among melanoma patients, the pretreatment levels of circulating exosomal PD-L1 were found to be negatively correlated with disease response to pembrolizumab. Importantly, a higher level of circulating exosomal PD-L1 before the treatment was associated with poorer clinical outcomes.69
There are also preclinical studies that indicate the removal of exosomal PD-L1 can increase the response rate of the anti-PD-L1 blockade, supporting the critical impact of exosomal PD-L1 in therapeutic resistance to anti-PD-L1 antibody treatment. In the mouse 4T1 breast tumor model, it was shown that the elimination of exosomal PD-L1 by inhibiting exosome secretion using knockdown of Rab27a in tumor cells considerably improved the efficiency of anti-PD-1 therapy and suppressed the tumor growth.70 The TRAMP-C2 model is a syngeneic model of prostate cancer81 that, like human prostate cancer, is resistant to anti-PD-L1 blockade.82 Removal of tumor exosomal PD-L1 from TRAMP-C2 cells using genetic mutations could efficiently enhance the effect of PD-L1 blockade, which included inhibition of tumor growth, elevated cellularity of the spleen, and the activation of a T cell response in lymph nodes with similar effects on the various activation, exhaustion, and proliferation markers. Interestingly, the injection of in vitro collected exosomes carrying PD-L1 was found to reverse all these outcomes in the TRAMP-C2 model, whereas, in the absence of PD-L1, the effect of the exogenously introduced exosomes was very low.82 Similarly, the removal of exosomal PD-L1 in the colorectal MC38 model inhibited tumor growth and extended survival through PD-L1 blockade treatment.73 Mechanistically, it was suggested that high levels of exosomal PD-L1 might reflect the “exhaustion” of T cells to a stage at which they can no longer be reinvigorated by anti-PD-1 therapy.73
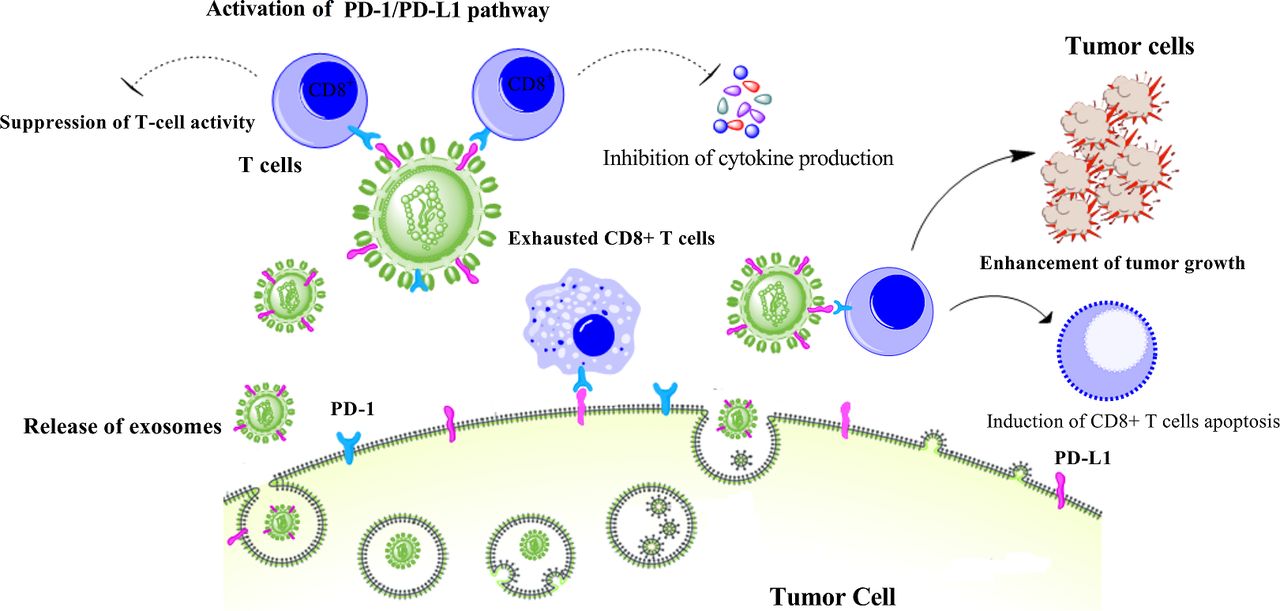
These findings are all consistent with exosomal PD-L1-mediated resistance to current anti-PD-L1/PD-1 therapies. However, the mechanism underlying the therapeutic resistance of exosomal PD-L1 is still unknown. One possible mechanism is that the delivered anti-PD-L1 antibodies can directly bind to exosomal PD-L1, and thereby few antibodies remain to inhibit PD-L1 on the surface of tumor cells. In addition, high levels of exosomal PD-L1 can compete with the administered anti-PD-L1/PD-1 antibodies (figure 2). Likewise, exosomes may reach targets that are sequestered from the antibody’s effect. Therefore, combination therapy using anti-exosomal treatment may synergize the therapeutic efficiency of anti-PD-1/PD-L1 blockade.

{kind=link}
{kind=link}
Exosomal PD-L1 induces acquired resistance to anti-PD-1/PD-L1 therapy. Exosomes carrying PD-L1 limit effectiveness of anti-PD-1/PD-L1 therapy through binding to antibodies and suppression of T-cell activity. However, elimination of PD-L1 exosomes can improve anti-PD-1/PD-L1 therapy. IFN-γ, interferon-gamma; PD-1, programmed cell death protein 1; PD-L1, programmed death-ligand 1.
Exosome elimination as an add-on therapy enhancing potency of anti-PD-1/PD-L1 antibodies
There is preclinical evidence that shows loss of PD-L1 expression from primary tumors along with the elimination of exosome secretion via pharmacological intervention or genetic manipulation mitigate the metastatic burden and elevate overall survival in a variety of tumor-bearing mice.69 70 73 Combination treatment with GW4869, an inhibitor of exosome secretion, and anti-PD-L1 mAb demonstrated the highest reduction in primary tumor burden in mice bearing 4T-1 tumor, showing a synergistic association between immune checkpoint inhibitors and exosome elimination.70 Supporting this, anti-PD-L1 efficacy was markedly enhanced in mice harboring tetracycline-inducible Rab27 knockdown 4T1 cells.70 Of equal relevance, an Rab27−/− MC38 cell line dramatically enlarged overall survival in mice receiving anti-PD-L1 mAb compared with either monotherapy alone.73 Similar results were also observed by an independent group in other tumor models.69 Altogether, these findings suggest that exosome elimination may act as an effective add-on therapy to improve the therapeutic efficiency of anti-PD-1/PD-L1 blockade in patients with cancer. Exosomes can be eliminated through suppressing their generation and secretion from tumor cells or removal of circulating exosomes from the bloodstream using haemofiltration.
Pharmacological approaches for suppressing exosome generation and secretion
Elimination of circulating exosomes has emerged as a novel and potentially useful therapeutic strategy for the development of anticancer drugs.83 Many reports have already shown that the reduction of exosome secretion or secreted exosomes, achieved by using a chemical inhibitor,84 85 genetic manipulation,86 or antibody,87 can suppress cancer metastasis and improve the efficiency of cancer chemotherapy. The effective strategies for exosome suppression are the use of pharmacological agents that reduce the level of exosomes by targeting different molecules involving in the generation, packaging, and release of them. However, since exosomes are implicated in intercellular communications and maintaining normal cellular physiology, this represents the most important limitations to be used as a therapeutic strategy, due to the potential toxicity and other side-effects caused by any partial or temporary inhibition of exosome secretion from normal cells when a drug candidate inhibits the secretion of exosomes from cancer cells. Using exosome inhibitors developed by drug repurposing strategy, a process of finding new indications for existing FDA-approved drugs, can reduce concerns about safety/toxicity, because these agents have already passed toxicity and safety tests in humans.85 88 For example, FDA-approved drugs, dimethyl amiloride (DMA) and omeprazole reduce exosome secretion through targeting H+/Na+ and Na+/Ca2+ channels89 that are known to involve in exosome release.90 DMA has been found to reduce the production of exosomes and eliminate their immune suppressive effects in tumor-bearing hosts and enhance the antitumor efficacy of the cyclophosphamide as a chemotherapeutic drug.89 Another possible option for inhibiting exosome secretion is the use of proton pump inhibitors (PPIs), which are widely prescribed for mitigating gastric acid.91 Vacuolar H+-ATPases can regulate pH of extracellular microenvironments and involve in tumor progression. PPIs through inhibiting vacuolar H+-ATPase-driven efflux pumps impair the release of acidic vesicles and vesicle-like structures by tumor cells and enhance the efficacy of chemotherapeutic agents.91 92 In addition, ceramide is known to involve in biogenesis and release of exosomes, and sphingomyelinases, enzymes promoting ceramide synthesis, participate in sorting the cargo as well. Notably, cell treatment with sphingomyelinase inhibitors was found to markedly reduce the release of exosomes.93 Moreover, sulfisoxazole, an FDA-approved oral antibiotic, was also identified as an inhibitor of exosome secretion from breast cancer cells through interference with endothelin receptor A, a member of G protein-coupled receptor (GPCR) family that plays a critical role in the biogenesis and secretion of exosome in breast cancer cells.88 Of note, sulfisoxazole exhibited significant antitumor and anti-metastatic impacts in mouse models of breast cancer xenografts, the decreased expression of proteins participated in exosome biogenesis and secretion, and induced lysosomal degradation of multivesicular endosomes.88 Interestingly, a quantitative high throughput screen assay of many pharmacologically active compounds approved for clinical application uncovers the lead compounds tipifarnib, neticonazole, climbazole, ketoconazole, and triademenol that inhibit exosome biogenesis and/or release by aggressive prostate cancer calls.85 Hence, utility of drug-repurposing for preventing exosome secretion may provide a useful and safe approach for unmasking the inhibitory effects of exosomal PD-L1 on immune function and therapeutic responses in cancer.
Exosome removal using extracorporeal haemofiltration
Another alternative promising approach is exosome removal from the blood circulation by extracorporeal haemofiltration, which would not represent the possible drug toxicity or interactions, thereby suggesting an advantage over pharmacological strategies. Aethlon ADAPT (adaptive dialysis-like affinity platform technology) therapy is a therapeutic haemofiltration approach with a mechanism of action similar to kidney dialysis or continuous renal replacement therapy method. In this approach, plasma and blood cells pass through the hollow‐fiber dialysis cartridges (<200 nm pore) that are packed with the immobilized affinity matrix. The component of matrices can be customized with our purpose and can be composed of different affinity agents including monoclonal antibodies, aptamers, and various ligands to specifically capture and remove soluble proteins from the plasma. During the circulation of plasma over the cartridge, target molecules can selectively interact with the matrix and be isolated from other molecules, while unbound components and blood cells of serum pass through the device.
The safety and efficacy of the Aethlon haemopurifier device have been evaluated in clearing the hepatitis C virus (HCV) from the blood of HCV-positive end-stage renal disease patients by using lectin affinity plasmapheresis.94 Of note, ADAPT therapy can be also used for targeting and removal of exosomes as an adjuvant treatment in cancer therapy. Marleau et al showed that haemopurifier technique can be employed for capturing tumor-derived exosomes based on specific molecules on the exosomal surface, such as high mannose exosomes to remove by lectins or HER2 positive exosomes to remove with anti-HER2 antibodies.95 Hence, it can be proposed that PD-L1 positive exosomes can be eliminated by Aethlon haemopurifier device equipped with a matrix containing anti-PD-L1 antibody, suggesting an adjunct therapeutic candidate to the standard of care cancer treatments. However, considering the role of circulating exosomal PD-L1 on the mediation of systemic immunosuppression, there are concerns about possible severe irAEs along with their elimination. Therefore, although some preclinical studies showed improved survival with no significant adverse effect in exosomal PD-L1 depleted mice bearing tumors,73 further preclinical toxicity testing is important to address the safety of exosomal PD-L1 elimination.
Conclusion and future perspectives
This review highlights and summarizes current knowledge regarding the role of exosomal PD-L1 in acquired resistance to anti-PD-1/PD-L1 therapy of cancer. In brief, exosomal PD-L1 can reprint the effect of tumor cell-surface PD-L1 in cancer progression by enabling tumor cells to escape from antitumor immunity. Exosomes can also deliver PD-L1 from origin tumor cells to other cell types with low or no PD-L1 expression and thereby suppress systemic antitumor immunity and memory. Notably, plasma levels of exosomal PD-L1 are negatively correlated with the rate of response to anti-PD-1/PD-L1 therapy in patients with cancer. Of note, there is evidence that indicates the removal of exosomal PD-L1 can increase the response to the anti-PD-L1 blockade in animal models. These findings are all consistent with exosomal PD-L1-mediated resistance to current anti-PD-L1/PD-1 therapies. Although the mechanism underlying therapeutic resistance of exosomal PD-L1 is still unknown, some possible mechanisms are proposed. Importantly, circulating exosomal PD-L1 can interact with anti-PD-1/PD-L1 antibodies and limit their therapeutic efficiency. Moreover, high levels of exosomal PD-L1 can compete with the administered anti-PD-L1/PD-1 antibodies. It is also possible that exosomes attain targets that are sequestered from the antibody’s effect. Therefore, combination therapy using anti-exosomal treatment may synergize the therapeutic efficiency of anti-PD-1/PD-L1 blockade. However, since normal cell-secreted exosomes are critical for maintaining normal physiology, a drug candidate should selectively inhibit cancer-specific exosomes to decrease potential toxicity/side-effects. As an alternative approach, removal of secreted PD-L1+ exosomes from the bloodstream may also be an efficient strategy, of course, if toxicity studies verify its safety and the lack of irAEs.
Ethics statements
Patient consent for publication
References
Footnotes
ZY and MY contributed equally.
Correction notice This article has been corrected since it first published. The provenance and peer review statement has been included and Dr. Momtazi-Borojeni's affiliation has been corrected.
Contributors ZY and SC contributed to the conception and design of the work. ZY and MY were major contributors in writing the manuscript. TM and CZ performed the database search and wrote a part of the manuscript. SH and MRK prepared the table and designed figures. AAM-B read and approved the final manuscript.
Funding This study was supported by the Science and Technology Program of Guangzhou (No. 201904010043), National Natural Science Foundation (No. 81802892, No. 82072637), and Medical Scientific Research Foundation of Guangdong (No. A2019453).
Competing interests None declared.
Provenance and peer review Not commissioned; externally peer reviewed.