Article Text

Abstract
Background Despite the preclinical promise of CD40 and 4-1BB as immuno-oncology targets, clinical efforts evaluating CD40 and 4-1BB agonists as monotherapy have found limited success. DuoBody-CD40×4-1BB (GEN1042/BNT312) is a novel investigational Fc-inert bispecific antibody for dual targeting and conditional stimulation of CD40 and 4-1BB to enhance priming and reactivation of tumor-specific immunity in patients with cancer.
Methods Characterization of DuoBody-CD40×4-1BB in vitro was performed in a broad range of functional immune cell assays, including cell-based reporter assays, T-cell proliferation assays, mixed-lymphocyte reactions and tumor-infiltrating lymphocyte assays, as well as live-cell imaging. The in vivo activity of DuoBody-CD40×4-1BB was assessed in blood samples from patients with advanced solid tumors that were treated with DuoBody-CD40×4-1BB in the dose-escalation phase of the first-in-human clinical trial (NCT04083599).
Results DuoBody-CD40×4-1BB exhibited conditional CD40 and 4-1BB agonist activity that was strictly dependent on crosslinking of both targets. Thereby, DuoBody-CD40×4-1BB strengthened the dendritic cell (DC)/T-cell immunological synapse, induced DC maturation, enhanced T-cell proliferation and effector functions in vitro and enhanced expansion of patient-derived tumor-infiltrating lymphocytes ex vivo. The addition of PD-1 blocking antibodies resulted in potentiation of T-cell activation and effector functions in vitro compared with either monotherapy, providing combination rationale. Furthermore, in a first-in-human clinical trial, DuoBody-CD40×4-1BB mediated clear immune modulation of peripheral antigen presenting cells and T cells in patients with advanced solid tumors.
Conclusion DuoBody-CD40×4-1BB is capable of enhancing antitumor immunity by modulating DC and T-cell functions and shows biological activity in patients with advanced solid tumors. These findings demonstrate that targeting of these two pathways with an Fc-inert bispecific antibody may be an efficacious approach to (re)activate tumor-specific immunity and support the clinical investigation of DuoBody-CD40×4-1BB for the treatment of cancer.
- immunotherapy
- T-lymphocytes
- dendritic cells
Data availability statement
Data are available on reasonable request. The datasets generated during and/or analyzed during the current study are available from the corresponding author on reasonable request.
This is an open access article distributed in accordance with the Creative Commons Attribution Non Commercial (CC BY-NC 4.0) license, which permits others to distribute, remix, adapt, build upon this work non-commercially, and license their derivative works on different terms, provided the original work is properly cited, appropriate credit is given, any changes made indicated, and the use is non-commercial. See http://creativecommons.org/licenses/by-nc/4.0/.
Statistics from Altmetric.com
Introduction
The unprecedented clinical success of immune checkpoint inhibitors (CPI) has demonstrated the potential for reactivation of antitumor immunity in cancer treatment. While CPI have revolutionized the treatment paradigm and prognosis for a subset of patients with advanced solid tumors, additional (combination) immunotherapies are needed to address unmet need as many patients progress after an initial response or experience primary resistance.1 2 In this context, targeting tumor necrosis factor receptor (TNFR) superfamily members, which deliver essential costimulatory activity for the generation of effective immune responses, has gained much attention.
4-1BB (CD137) is a costimulatory TNFR expressed on immune cells, including T cells, natural killer (NK) cells and monocytes.3 4 While naïve T cells lack 4-1BB expression, the receptor is upregulated on T-cell receptor (TCR) engagement. Crosslinking of 4-1BB activates the MAPK and NF-κB signaling pathways and enhances T-cell proliferation and cytokine production (including interferon (IFN)-γ and tumor necrosis factor (TNF)-α), enhances cytotoxic T-cell effector functions and prevents activation-induced cell death of T cells.3 4 In preclinical studies, treatment with agonistic monoclonal antibodies (mAbs) directed against 4-1BB leads to tumor regression and protects against secondary tumor challenge by augmenting T-cell and NK-cell mediated anti-tumor immunity.3 4
CD40 is a stimulatory receptor that is primarily expressed on antigen presenting cells (APCs), including dendritic cells (DCs), B cells, monocytes and macrophages.5 6 Therefore, unlike other TNFR family members that are primarily expressed on T cells, engagement of CD40 leads to activation and maturation of the CD40-expressing APCs. CD40 ligand (CD40L) expression on immune cells is mainly localized to lymphocyte subsets such as T cells, NK cells and B cells, but is most prominent on activated CD4+ T cells.5 6 DCs that have been activated through CD40 stimulation upregulate the costimulatory molecules CD80 and CD86 as well as ligands of other TNFRs and secrete enhanced levels of immunostimulatory cytokines such as interleukin (IL)-12 and TNF-α.7 Owing to this central role in the activation of adaptive immune responses, CD40 is an attractive target for augmentation of antitumor immune responses.8 Importantly, by directly binding to DCs, CD40 agonistic agents can substitute for DC activation via CD40L+ CD4+ T cells, thereby leading to the generation of robust CD8+ T-cell mediated antitumor immunity independent of CD4+ T-cell help.9–11
The use of agonistic antibodies to induce CD40 immunostimulatory signaling in APCs and to enhance activation of 4-1BB-expressing T cells, respectively, may provide opportunities to boost antitumor immune responses within the tumor and in tumor-associated lymphoid tissue. In preclinical studies, bispecific antibodies that activate 4-1BB through crosslinking with a tumor-associated secondary target enhanced the antitumor immune response in the tumor microenvironment (TME).12–15 Additional costimulation of CD40 could further enhance costimulatory signaling provided by APCs, resulting in enhanced priming and (re)activation of tumor-specific immunity.
While these preclinical findings have validated CD40 and 4-1BB as targets for cancer immunotherapy, clinical efforts using agonistic single targeted agents have met limited success. Modest monotherapy response rates have been recorded in several clinical trials of CD40-agonistic mAbs, despite clear evidence of pharmacodynamic activity.6 16–18 4-1BB agonists have either shown limited efficacy at well-tolerated doses or hepatotoxicity at efficacious doses.4 19 20 The underlying biology of these pathways provides a clear rationale for the combined bispecific targeting of 4-1BB and CD40 to enhance response rates and broaden the therapeutic window.
We hypothesized that dual targeting and conditional stimulation of 4-1BB and CD40 using an Fc-inert bispecific antibody may combine these two complementary mechanisms to further enhance antitumor immunity. Here, we describe the generation and preclinical characterization of DuoBody-CD40×4-1BB (GEN1042/BNT312), a novel investigational bispecific agonistic antibody that combines targeting of costimulatory molecules CD40 and 4-1BB. Preclinical proof-of-concept data are presented in addition to initial pharmacodynamics results from the dose-escalation phase of our first-in-human clinical trial.
Materials and methods
Antibodies
Cell lines, commercially available antibodies and research antibodies produced at Genmab are listed in online supplemental methods. Monoclonal antibodies (mAbs) were recombinantly produced in a human IgG1 backbone, using previously described methods.21 The mutations L234F, L235E and D265A were introduced to abrogate Fc binding to IgG Fc receptors (FcγR) and complement component C1q.22 The F405L and K409R DuoBody mutations were introduced to allow generation of bsAbs by controlled Fab-arm exchange.23 24 The HIV-1 gp120-specific isotype control antibody (mAb-ctrl) based on IgG1-b1225 was used to generate bsAbs with one non-binding Fab arm (eg, bsAb-ctrlx4-1BB). The detailed procedure for the generation and selection of mAb-CD40-FEAL, mAb-4-1BB-FEAR and DuoBody-CD40×4-1BB is described in online supplemental methods.
Supplemental material
Cell isolation and enrichment
Peripheral blood mononuclear cells (PBMCs) were isolated from buffy coats of healthy human blood donors (Transfusionszentrale, University Hospital, Mainz, Germany) by density centrifugation over a Ficoll-Paque gradient (VWR, Ficoll-Paque PLUS medium, 17-5446-02), according to the manufacturer’s instructions.
CD14+ monocytes were enriched from PBMCs of HLA-A*02-positive donors by magnetic-activated cell sorting (MACS), using CD14 magnetic microbeads (Miltenyi, 130-050-201), AutoMACS columns (Miltenyi, 130-021-101) and an AutoMACS Pro Separator (Miltenyi, 130-092-545), according to the manufacturer’s instructions. Similarly, CD8+ T-cell populations were enriched from the CD14- fraction or whole PBMCs by MACS using CD8 microbeads (Miltenyi, 130-045-201).
Differentiation of immature dendritic cells (iDC)
CD14+ monocytes were either isolated as described above or obtained from Precision for Medicine (81 400–5M) or BioIVT. For differentiation into iDC, CD14+ cells were incubated with 100 ng/mL GM-CSF (eg, Biolegend, 766106) and 50–300 ng/mL IL-4 (eg, Biolegend, 766206) in suspension culture flasks in DC or MLR medium (online supplemental methods). Fresh medium supplemented with GM-CSF and IL-4 was supplied as appropriate. Non-adherent and adherent cells were harvested between days 5 and 7, pooled and collectively referred to as iDCs.
Electroporation
iDC or CD8+ T cells were suspended at a concentration of 20×106 or 60×106 cells/mL in X-Vivo15 medium, respectively. For each electroporation, a 250 μL cell suspension was transferred to electroporation cuvettes (4.0 mm gap size, VWR International GmbH, 732-0023) and kept at RT (iDC) or on ice (T cells). RNA encoding claudin 6 (CLDN6; 0.3–3 μg) was electroporated into iDCs (300 V, 12 ms, 1 pulse) using an ECM 830 Electroporation System (BTX, 45-0002). RNA encoding the alpha and beta chains of the CLDN6 TCR (HLA-A2/CLDN6-TCR; 10 μg each) was electroporated into T cells (500 V, 3 ms, 1 pulse). Immediately after electroporation, 1 mL of prewarmed assay medium (online supplemental methods) was added. The iDC suspension was transferred to a 6-well plate (Greiner Bio-One GmbH, 657-160) and incubated O/N. The T cells were transferred to a 15 mL tube and incubated for 1 h. Expression of the electroporated proteins was confirmed by flow cytometry.
Imaging of DC:T-cell interaction
DCs were seeded on poly-L-lysine (0.1 µg/mL; 2×106 cells/well) coated coverslips for 4 hours, followed by coculture with anti-CD3/anti-CD28 prestimulated CD8+ T cells (4×106 cells/well) in the presence of 0.5 µg/mL of DuoBody-CD40×4-1BB or control antibodies and further analyzed by immunofluorescence.
Claudin 6 (CLDN6)-electroporated iDC were seeded in a 96-well plate (1×104 cells/well). After O/N incubation, CLDN6-TCR-electroporated CD8+ T cells were added (2×104 cells/well) and the cells cocultured in the presence of 0.125 µg/mL of DuoBody-CD40×4-1BB or control antibodies and further analyzed by live-cell imaging.
Reporter assays
CD40 agonist activity was evaluated using human CD40 effector cells (T&U GloResponse NFkB-luc2P/U2OS cells; Promega, CS1979A06) and the BioGlo Luciferase Assay (Promega, G720A). CD40 effector cells were cocultured with K562 human chronic myelogenous leukemia cells, which were stably transfected with human 4-1BB (K562_h4-1BB) or without K562_h4-1BB cells, in the presence of antibodies. 4-1BB agonist activity was evaluated using human 4-1BB effector cells (T&U GloResponse NFkB-luc2/4-1BB Jurkat cells; Promega, CS196003) and the BioGlo Luciferase Assay. 4-1BB effector cells were cocultured with K562 cells, which were stably transfected with human CD40 (K562_hCD40), or without K562_hCD40 cells, in the presence of antibodies. Luminescence was recorded using an Infinite F200 Pro plate reader (Tecan Life Sciences) or an EnVision multilabel plate reader (EnVision).
B-cell activation
CD19+ B cells were isolated (enriched) from buffy coats by MACS using CD19 magnetic microbeads (Miltenyi Biotec GmbH, cat. no. 130-050-301) and were collected in prewarmed B-cell medium. B cells were cocultured with K562 cells or K562_h4-1BB cells at a 1:1 ratio (2.5×105 cells/mL for both cell types) in the presence of DuoBody-CD40×4-1BB at 37°C, 5% CO2 for 2 days. Expression of activation markers CD69 and CD86 was evaluated by flow cytometry.
T-cell proliferation assays
PBMC were labeled with carboxyfluorescein succinimidyl ester (CFSE) using the Vybrant CFDA SE Cell tracer kit (Life Technologies GmbH, V12883), and stimulated with an anti-CD3 antibody in T-cell assay media at 7.5×104 cells per well in round-bottom 96-well plates (Costar, 734–1797). The cells were cultured in the presence of DuoBody-CD40×4-1BB or control antibodies for 4 days, and CFSE dilution was analyzed by flow cytometry. Supernatants were harvested for cytokine analysis 48 hours after start of the culture.
CD8+ T cells were isolated from healthy donor PBMCs (HLA-A*02) and electroporated with RNA encoding an HLA-A*02-restricted, CLDN6-TCR. Cells were labeled with CFSE or alternative CellTrace proliferation dyes as described above and cocultured with CLDN6-electroporated autologous iDC in round-bottom 96-well plates (5×104 T cells and 5×103 iDC per well). Cells were treated with DuoBody-CD40×4-1BB or control antibodies for 4 days. Supernatants were harvested for cytokine analysis, and cells were analyzed for T-cell proliferation by flow cytometry as above.
iDC maturation and mixed-lymphocyte reaction (MLR) assays
To obtain mature DCs (mDCs), the iDCs were incubated with 1×LPS (ThermoFisher, 00-4976-93) for 24 hours prior to initiation of the MLR. CD8+ T cells were incubated in MLR medium with 10 ng/mL IL-2 (BioLegend, 589106) O/N, prior to initiation of the MLR.
CD8+ T cells were cocultured with either iDCs or mDCs (2×105 T cells: 2×104 DC per well) in AIM-V medium and incubated with DuoBody-CD40×4-1BB (monotherapy (iDC:CD8+ T cell and mDC:CD8+ T-cell cocultures) or concurrent treatment with pembrolizumab (mDC:CD8+ T-cell cocultures; Research-grade, Selleckchem, A2005)) or control antibodies for 5 days. Supernatants were harvested for cytokine analysis.
Tumor-infiltrating lymphocyte (TIL) assay
Fresh tumor resections were obtained from cancer patients (all patients provided written informed consent), collected in transport medium (Supplementary Methods) and kept on ice until further processing. Specimens were washed three times in wash medium (online supplemental methods), cut into pieces of about 1–2 mm3 and used for TIL expansion. The pieces were transferred to a 24-well plate (VWR international, 734–1604) at two pieces per well in 1 mL TIL medium (Supplementary Methods) and incubated with DuoBody-CD40×4-1BB or control antibodies in the presence of 10 U/mL IL-2. Fresh medium supplemented with antibody was supplied as appropriate. After 10–15 days, TIL numbers were analyzed by flow cytometry and quantified using reference beads. In addition, TILs pooled from individual wells by treatment condition were subjected to TCR sequencing.
Clinical study
In the ongoing first-in-human clinical study (registered on https://www.clinicaltrials.gov by identifier NCT04083599) escalating doses of DuoBody-CD40×4-1BB were administered intravenously.
Statistical analysis
Statistical significance was determined using Mann-Whitney U test for pairwise comparisons and Friedman test with Dunn’s multiple comparisons test or one-way analysis of variance with Dunnett’s multiple comparisons test, as indicated in the figure legends.
Results
Generation of DuoBody-CD40x4-1BB and target binding characteristics
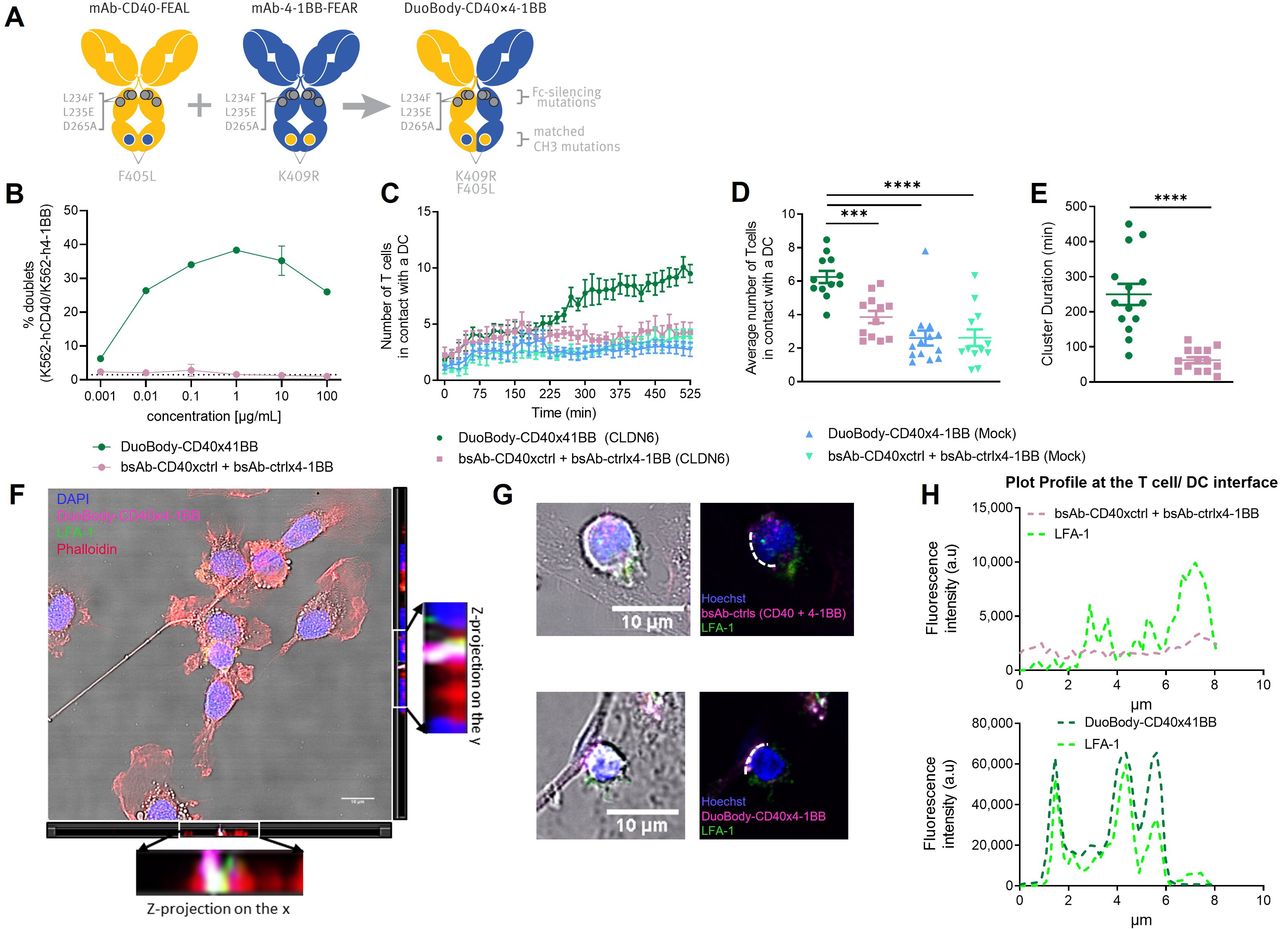
To evaluate the immunostimulatory potential of targeting CD40 and 4-1BB with a bsAb, we generated DuoBody-CD40×4-1BB by controlled Fab-arm exchange (figure 1A). The L234F, L235E and D265A mutations, which have previously been shown to limit binding to FcγRI, FcγRII and FcγRIII and complement component C1q,22 were introduced to avoid CD40 or 4-1BB clustering and activation through C1q-mediated and FcγR-mediated crosslinking of target-bound antibody. We confirmed lack of binding of DuoBody-CD40×4-1BB to FcγRIIa, FcγRIIb and FcγRIII (online supplemental figure S1A). Thereby, the bsAb was designed to induce 4-1BB agonist activity exclusively on simultaneous binding to CD40 and vice versa.

DuoBody-CD40×4-1BB reinforces the immunological synapse through target crosslinking. (A) Generation of DuoBody-CD40×4-1BB by cFAE of Fc-silenced mAb-CD40 and mAb-4-1BB. The parental antibodies contain matched mutations in the CH3 domain (F405L/K409R) that drive heterodimerization of the Fab arms and formation of bispecific molecules during cFAE, as well as Fc-silencing mutations (L234F, L235E and D265A) that abrogate binding to FcγR and C1q. (B) Simultaneous binding of DuoBody-CD40×4-1BB to human CD40-transgenic K562 cells labeled with CellTrace Violet and human 4-1BB-transgenic K562 cells labeled with CellTrace FarRed was analyzed by flow cytometry. Double-positive doublets were quantified as percentage within the live cell population (mean±SD; triplicate wells). (C–E) T cells electroporated with a CLDN6-specific TCR were cocultured with CLDN6-electroporated or mock-electroporated autologous iDCs in the presence of 0.125 µg/mL DuoBody-CD40×4-1BB or control antibodies, and T-cell/iDC clusters were visualized over time by live-cell imaging. Quantification of the number of T cells in contact with a given DC over time (C), or on average (D), as well as the duration of these DC/T-cell clusters (E) is shown. Error bars represent SD. ****P<0.0001; ***p<0.001; one-way ANOVA with Dunnett’s multiple comparisons test (D). ****P<0.0001; Mann-Whitney test (E). (F) Single Z plane of merged brightfield and fluorescent image of iDC cocultured with preactivated CD8+ T cells in the presence of Alexa Fluor 647-conjugated DuoBody-CD40×4-1BB (magenta)and LFA-1 (in green) antibodies, on the x and y axes the z-stack of the same picture with the relative zoom in. Nuclei were counterstained with Hoechst (in blue). (G) Representative fluorescent images of cocultures as in (E), in the presence of DuoBody-CD40×4-1BB or control antibodies. The white dashed line represents the interface between the DC and T cell. Scale bars: 10 µm. (H) LUT profile of LFA-1 and DuoBody-CD40×4-1BB or the combination of bsAb-CD40xctrl and bsAb-ctrlx4-1BB at the DC-T-cell interface, indicated by the white dashed line. ANOVA, analysis of variance; cFAE, controlled Fab-arm exchange; iDC, immature dendritic cell; LFA-1, leukocyte functional antigen 1; LUT, look-up table.
DuoBody-CD40×4-1BB bound to recombinant human CD40 and 4-1BB with affinities of 1.0 nM and 0.17 nM, respectively (online supplemental table S1 and figure S1B,C). The CD40-specific and 4-1BB-specific Fab arms of DuoBody-CD40×4-1BB showed dose-dependent binding to primary human B cells and activated T cells, respectively (online supplemental figure S1D,E). Binding to cell membrane-associated targets was confirmed using cell lines transfected with either human CD40 or human 4-1BB (online supplemental figure S1F,G).
DuoBody-CD40x4-1BB reinforces the immunological synapse
As DuoBody-CD40×4-1BB efficiently bound CD40 and 4-1BB individually, we next addressed the question whether it could promote cell-cell interactions by simultaneously binding both targets. To this end, we analyzed cell-cell interactions in the presence of DuoBody-CD40×4-1BB or monovalent control bsAbs comprised of a CD40 or 4-1BB binding Fab arm and a non-binding Fab arm (bsAb-CD40xctrl and bsAb-ctrlx4-1BB). DuoBody-CD40×4-1BB fostered the formation of doublets of CD40-positive and 4-1BB-positive cells at a broad range of concentrations, with maximal frequency of doublets between 0.1 and 10 µg/mL. The combination of monovalent antibodies did not induce any doublet formation (figure 1B).
The effect of DuoBody-CD40×4-1BB on interactions between monocyte-derived DCs and T cells was visualized using fluorescence microscopy, using claudin 6 (CLDN6) as a model antigen. We cocultured CD8+ T cells electroporated with a CLDN6-specific TCR with CLDN6-electroporated autologous DCs and assessed DC/T-cell interactions over time by live-cell imaging. DuoBody-CD40×4-1BB significantly promoted the formation of DC/T-cell clusters and prolonged the duration of the DC/T-cell interactions, compared with the combination of monovalent control bsAbs (figure 1C–E). Importantly, the effect on cluster formation required TCR recognition of cognate antigen, as it was not observed when mock-electroporated DCs were used.
We confirmed enhanced DC/T-cell interactions in the presence of DuoBody-CD40×4-1BB using immunofluorescence in a modified experimental setup, where CD8+ T cells were preactivated by stimulation with anti-CD3/anti-CD28 coated microbeads before coculture with DCs (online supplemental figure S2A). DuoBody-CD40×4-1BB induced significantly higher frequencies of DC/T-cell interactions compared with monovalent control bsAbs or a combination thereof (online supplemental figure S2B,C). Further analyses showed that DuoBody-CD40×4-1BB preferentially localized to the DC/T-cell interface, whereas the monovalent control bsAbs showed a less focused localization pattern on the cell surface (figure 1F–G). In addition, DuoBody-CD40×4-1BB colocalized with leukocyte functional antigen 1 (LFA-1), a marker of the immunological synapse between T cells and APCs,26 whereas colocalization of control antibodies with LFA-1 was limited (figure 1F–H and online supplemental figure S2D,E). Taken together, these results showed that DuoBody-CD40×4-1BB reinforces the immunological synapse by crosslinking both targets, thereby enhancing interactions between DCs and T cells.

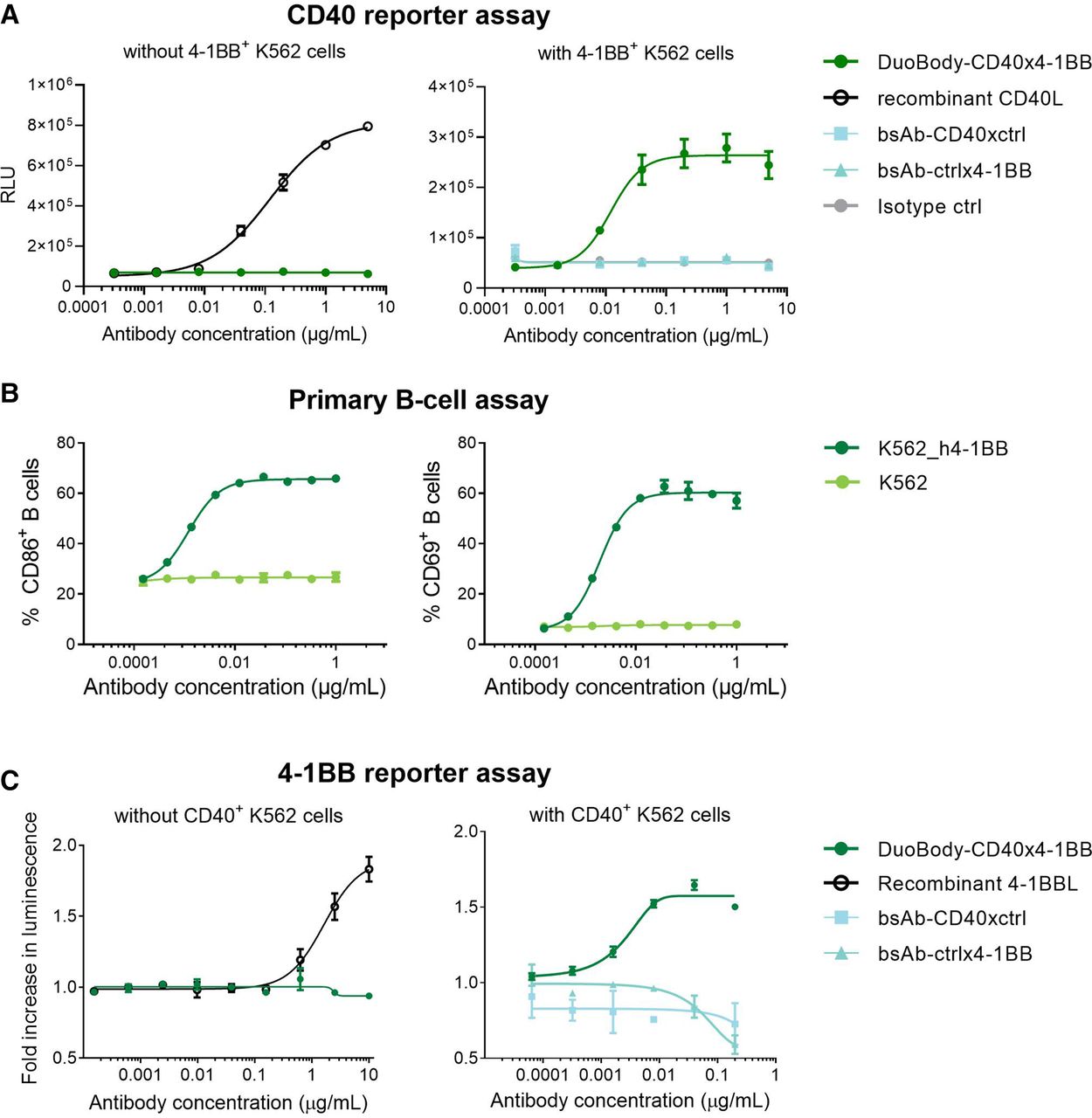
DuoBody-CD40×4-1BB exhibits conditional agonist activity. (A) CD40 bioluminescent cell-based reporter assay in the absence or presence of 4-1BB-expressing K562 cells. Cells were incubated with DuoBody-CD40×4-1BB, recombinant CD40L or control antibodies. (B) Human peripheral blood B cells were cocultured with 4-1BB-transgenic or wild type K562 cells in the presence of DuoBody-CD40×4-1BB for 48 hours. Expression of CD69 and CD86 on CD20+ B cells was analyzed by flow cytometry. Data shown are mean percentages of CD69+ or CD86+ B cells from one representative donor (n=3). Error bars represent SD. (C) 4-1BB bioluminescent cell-based reporter assay in the absence or presence of CD40-expressing K562 cells. Fold luminescence-induction of cells incubated with DuoBody-CD40×4-1BB or control antibodies, compared with treatment control (isotype control antibody or media only) is shown. Error bars represent SD. RLU, relative luminescence units.
DuoBody-CD40x4-1BB exhibits conditional agonist activity
Next, we performed studies using cell-based reporter assays to evaluate the effect of target crosslinking on receptor activation. We assessed CD40 agonist activity using a U2OS reporter cell line that expresses an NF-κB-driven luciferase reporter. In this cell line, DuoBody-CD40×4-1BB induced robust activation of reporter luminescence, whereas monospecific control bsAbs had no effect (figure 2A). Importantly, CD40 downstream signaling and reporter activation only occurred in coculture with 4-1BB-transgenic cells, demonstrating that CD40 agonist activity was conditional, that is, dependent on crosslinking to 4-1BB. Conditional CD40 agonist activity was confirmed in a primary human B-cell assay, where DuoBody-CD40×4-1BB dose-dependently increased the percentage of B cells expressing activation markers CD86+ and CD69+ only on coculture with 4-1BB-transduced cells (figure 2B). We next used 4-1BB-expressing Jurkat cells harboring an NF-κB reporter construct to investigate 4-1BB agonist activity. DuoBody-CD40×4-1BB induced reporter activity in the Jurkat cells only in the presence of CD40-expressing cells (figure 2C). As before, reporter activity in the presence of CD40-expressing cells was not observed when monospecific control bsAbs were used. These results show that DuoBody-CD40×4-1BB exhibits conditional CD40 and 4-1BB agonist activity that is dependent on crosslinking of both targets.
DuoBody-CD40x4-1BB induces DC maturation in vitro
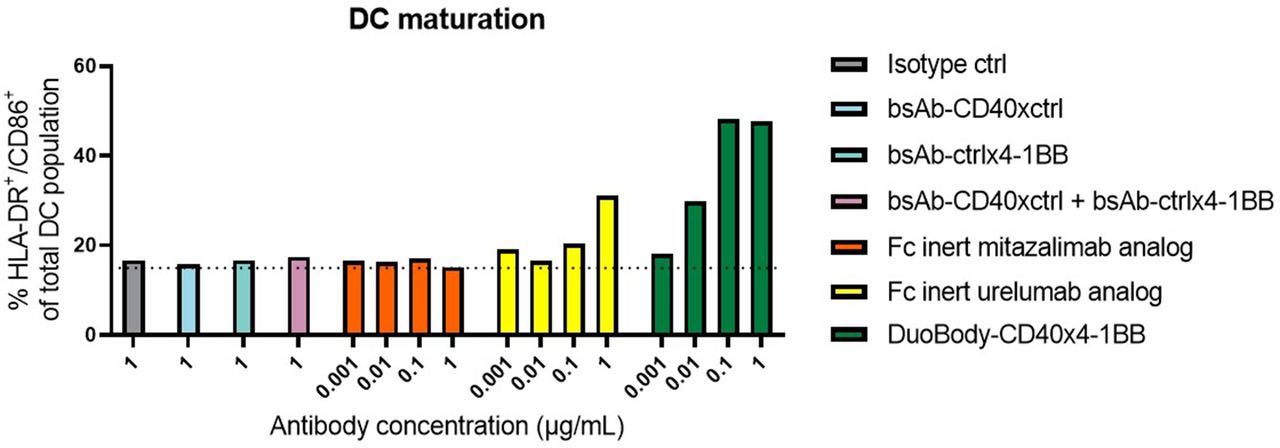
To further explore the immunostimulatory effect of DuoBody-CD40×4-1BB on a functional level, we analyzed DC maturation in MLR. To this end, we cocultured human monocyte-derived immature DCs (iDCs) with allogeneic naïve CD8+ T cells in the presence of DuoBody-CD40×4-1BB or control antibodies. DuoBody-CD40×4-1BB induced DC maturation at 0.01–10 µg/mL, as shown by the increased percentage of HLA-DR+/CD86+ DCs (figure 3). DC maturation induced by DuoBody-CD40×4-1BB was dependent on binding to both targets, as reflected by the lack of DC maturation induced by the combination of monovalent control bsAbs. Moreover, our results demonstrate that DuoBody-CD40×4-1BB, which has a functionally inert Fc domain, functions independent of FcγR mediated crosslinking. By contrast, Fc-inert analogs of clinical mAbs mitazalimab (targeting CD40) or urelumab (targeting 4-1BB) induced no or limited DC maturation.

DuoBody-CD40×4-1BB induces DC maturation. Immature DCs from healthy donors were cocultured with purified, allogeneic CD8+ T cells in the presence of DuoBody-CD40×4-1BB, Fc-inert analogs of clinical mAbs mitazalimab and urelumab (0.001–1 µg/mL) or control antibodies (1 µg/mL; for combination of control antibodies 1 µg/mL of both antibodies was added) for 5 days. The percentage of HLA-DR+CD86+ DCs was measured by flow cytometry. Data from one donor are shown. Dotted line shows percentage of HLA-DR+CD86+ DCs in DC-T-cell cultures in the absence of treatment. DC, dendritic cell.
DuoBody-CD40x4-1BB enhances T-cell activation in vitro
We next addressed immunostimulation of T cells through 4-1BB agonist activity of DuoBody-CD40×4-1BB. T-cell proliferation was analyzed in anti-CD3 stimulated PBMCs (polyclonal assay) or cocultures of CLDN6-electroporated autologous iDCs and CLDN6-TCR+ CD8+ T cells (antigen-specific assay). DuoBody-CD40×4-1BB significantly enhanced proliferation of activated CD8+ T cells in both the polyclonal (figure 4A) and antigen-specific setting (figure 4B). The enhanced T-cell proliferation induced by DuoBody-CD40×4-1BB was conditional, as none of the monospecific CD40-targeting or 4-1BB-targeting antibodies, either alone or in combination, affected T-cell proliferation. Similarly, DuoBody-CD40×4-1BB also conditionally enhanced proliferation of anti-CD3 stimulated CD4+ T cells, although to a lesser extent compared with CD8+ T cells (online supplemental figure S3A). Importantly, DuoBody-CD40×4-1BB specifically induced proliferation in activated T cells, as no effect on proliferation of CD8+ or CD4+ T cells was observed in the absence of anti-CD3 or cognate antigen stimulation. CD8+ T-cell proliferation induced by DuoBody-CD40×4-1BB was dose-dependent, with EC50 values in the picomolar range for both the polyclonal (124±49 pM; figure 4C) and the antigen-specific assay (55±11 pM; figure 4D).

DuoBody-CD40×4-1BB enhances T-cell activation in vitro. (A) CFSE-labeled human PBMCs were stimulated with 0.03 µg/mL anti-CD3 and incubated with DuoBody-CD40×4-1BB or control antibodies (0.2 µg/mL) for 4 days. CFSE dilution in CD8+ T cells was analyzed by flow cytometry. Expansion index of individual donors and mean±SD (n=7) is shown. (B) CD8+ T cells were electroporated with RNA encoding an HLA-A2/CLDN6-specific TCR, labeled with CFSE and cocultured with autologous DCs electroporated with CLDN6-encoding RNA in presence of the DuoBody-CD40×4-1BB or control antibodies (0.2 µg/mL; for combination of control antibodies 0.2 µg/mL of both antibodies was added) for 4 days. Proliferation was measured as described in (A). Expansion index of individual donors and mean±SD (n=9) is shown. ***P<0.001; *p<0.05; Friedman test. (C–D) Polyclonal (C) and antigen-specific T-cell proliferation assays (D) as described in A–B with increasing concentrations of DuoBody-CD40×4-1BB or isotype control antibody. Data shown are mean expansion index (±SD) of triplicate wells of one representative donor (n=12 for polyclonal assay, n=5 for antigen-specific assay). (E) Cytokine concentrations in supernatant taken after 48 hours from cultures as described in A–B. Data shown are mean concentration ±SD of triplicate wells from one representative donor (n=3 for polyclonal assay, n=5 for antigen-specific assay). (F–G) CD8+ T cells electroporated with a CLDN6-specific TCR were cocultured with CLDN6-expressing K562 cells for 48 hours. The percentage of IFN-γ+ cells or GzmB+CD107a+ cells (F), and expression levels of GzmB and CD107a normalized to the isotype control (G) were analyzed by flow cytometry. Data from individual donors and mean±SD (n=8) are shown. ***P<0.001; **p<0.01; One-way ANOVA with Dunnett’s multiple comparisons test (F) or paired t-test (G). ANOVA, analysis of variance; CFSE, carboxyfluorescein succinimidyl ester; DC, dendritic cell; IFN, interferon; PBMC, peripheral blood mononuclear cell.
We further explored the effect of DuoBody-CD40×4-1BB on T-cell effector functions, such as cytokine secretion and cytotoxic activity. In both the polyclonal and antigen-specific assays, DuoBody-CD40×4-1BB augmented the secretion of IFN-γ and TNF-α in a dose-dependent manner (figure 4E and online supplemental figure S3B). Increased cytokine secretion was again dependent on target crosslinking, as monospecific control antibodies did not lead to an increase even when combined. In MLRs of iDCs and CD8+ T cells, treatment with DuoBody-CD40×4-1BB also led to a substantial increase of IFN-γ, which was dependent on target crosslinking, increased compared with a mitazalimab analog and increased at lower concentrations of DuoBody-CD40×4-1BB compared with a urelumab analog (online supplemental figure S3C). Similarly, when LPS-matured DCs were used for the MLR, DuoBody-CD40×4-1BB treatment led to a dose-dependent increase of IFN-γ (online supplemental figure S3D).
The capacity of DuoBody-CD40×4-1BB to enhance CD8+ T-cell cytotoxic activity was evaluated by intracellular staining of cytotoxicity-associated molecules, using the CLDN6 model antigen and K562 cells stably expressing CD40. The percentage of CLDN6-specific CD8+ T cells expressing cytotoxic molecule GzmB and the degranulation marker CD107a was significantly increased after co-culture with CLDN6-electroporated K562_CD40 cells, along with a concurrent increase of IFN-γ+ cells (figure 4F). Expression of GzmB and CD107a was also increased on a per-cell level (figure 4G). Both the increase in frequency and intensity were dependent on simultaneous engagement of both targets, as the expression levels of GzmB and CD107a after exposure to a combination of monospecific control antibodies was comparable to the isotype control. Taken together, these findings demonstrated that DuoBody-CD40×4-1BB enhances T-cell activation, cytokine secretion and cytotoxicity through conditional 4-1BB agonist activity.
DuoBody-CD40x4-1BB enhances tumor-infiltrating lymphocyte expansion ex vivo
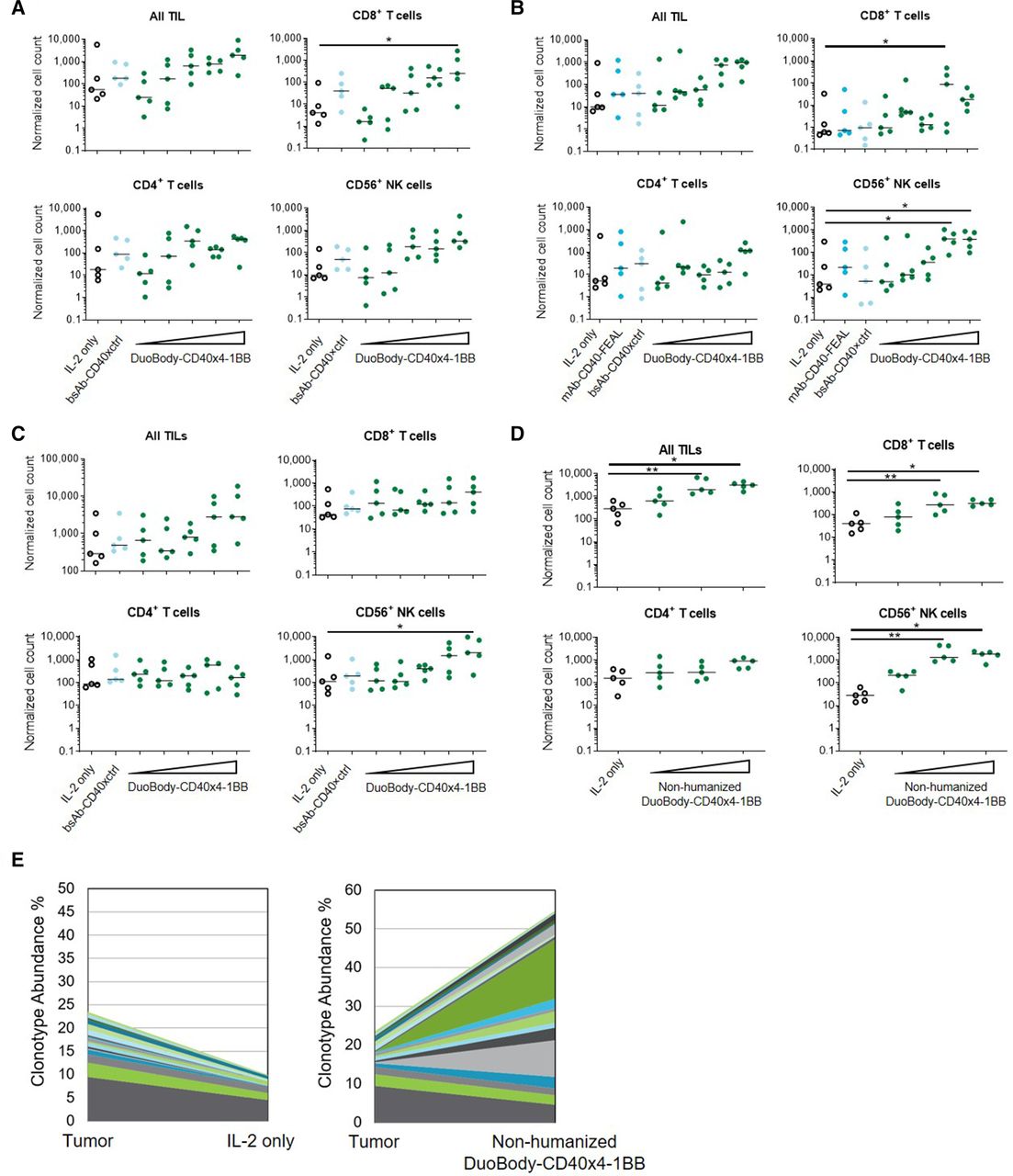
As DuoBody-CD40×4-1BB had a profound effect on the activation and function of healthy donor PBMC-derived T cells, we next addressed the question whether it could also improve the expansion of TILs from patient-derived tissues. To investigate this, we cultured fragments of tumor that had been resected from three patients with non-small cell lung cancer (NSCLC) and a patient with colon cancer in the presence of low-dose IL-2. In all cases, formation of TIL microclusters was observed in cultures treated with DuoBody-CD40×4-1BB (or a non-humanized DuoBody-CD40×4-1BB variant), but not control cultures, within 3–10 days after culture initiation (online supplemental figure S4A). Accordingly, flow cytometry analysis after 2 weeks of culture showed that DuoBody-CD40×4-1BB treatment led to a dose-dependent expansion of bulk TILs in all specimens (figure 5A–D). Numbers of CD8+ T cells were significantly increased within DuoBody-CD40×4-1BB-treated TILs at concentrations ≥0.05 µg/mL in one colon and two out of three NSCLC specimens, whereas on treatment with CD40 monospecific control antibodies changes were not significant. Furthermore, in the TIL cultures that were derived from the patients with NSCLC, NK cells were also significantly increased at concentrations ≥0.05 µg/mL. While there was a trend of increased CD4+ T-cell expansion in three out of four specimens, the difference was not statistically significant when compared with control cultures.

DuoBody-CD40×4-1BB enhances TIL expansion. Tumor tissues resected from patients were cut into pieces of 1–2 mm3 and cultured in the presence of IL-2 (10 U/mL) and DuoBody-CD40×4-1BB (0.0008, 0.003, 0.0125, 0.05 or 0.2 µg/mL), a non-humanized variant of DuoBody-CD40×4-1BB (0.01–1 µg/mL), control antibodies (0.2 µg/mL) or IL-2 only for 10–15 days. (A–D) Cell numbers after expansion were determined by flow cytometry, and normalized to reference beads. Total TIL, CD8+, CD4+ T-cell and NK-cell numbers are shown for a patient with colon cancer (A) and three patients with NSCLC (B–D) *p<0.05. One-way ANOVA with Dunnett’s multiple comparisons test. (E) TCR repertoire analysis was performed by TRB RNA sequencing of TIL expanded in (D), in the presence of a non-humanized variant of DuoBody-CD40×4-1BB (0.1 µg/mL) or IL-2 only. Cumulative frequency of shared clonotypes, the 20 most abundant clonotypes in the DuoBody-CD40×4-1BB-treated cultures are shown. ANOVA, analysis of variance; NK, natural killer; NSCLC, non-small cell lung cancer; TCR, T-cell receptor; TIL, tumor-infiltrating lymphocyte.
We performed sequencing of TCR β-chains to compare the abundance of individual TCR clonotypes in the expanded TILs and fragments in a specimen derived from one of the patients with NSCLC. TCR β sequencing showed specific expansion of T-cell clonotypes in TIL cultures treated with a non-humanized variant of DuoBody-CD40×4-1BB (figure 5E). T cells that were expanded by the non-humanized variant of DuoBody-CD40×4-1BB included clonotypes that were abundantly prevalent in fragments of the resected tissue, as well as initially underrepresented clonotypes (online supplemental figure S4B). Taken together, these data indicate that DuoBody-CD40×4-1BB is capable of inducing expansion of TILs from primary tumor tissue and may result in an enrichment of specific CD8+ T-cell clonotypes.
Combination of DuoBody-CD40x4-1BB with PD-1 blockade further amplifies the immune response in vitro
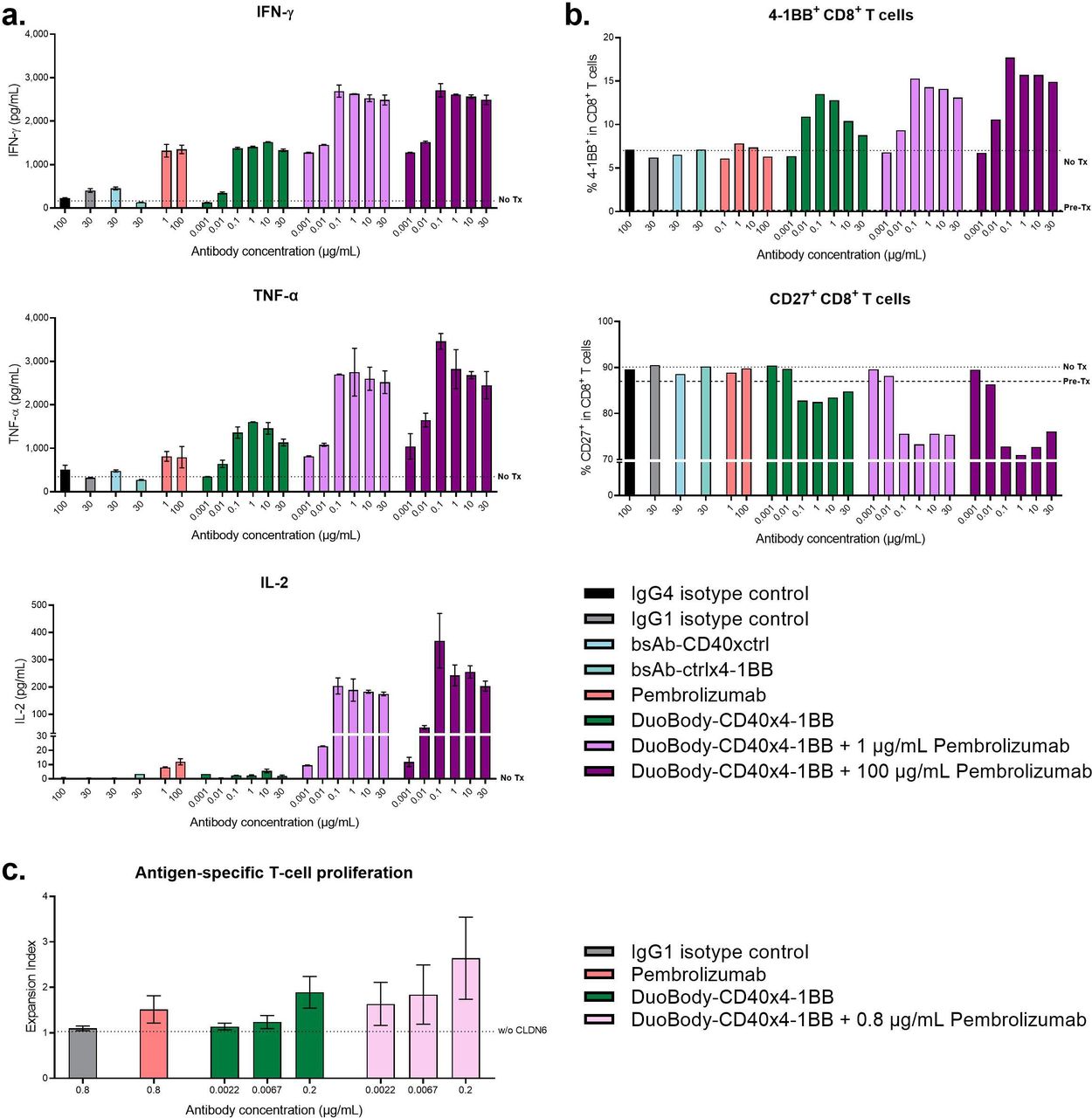
Blockade of PD-L1/PD-1 signals and conditional 4-1BB agonist activity may synergize to enhance T-cell effector functions and antitumor immunity. Using an mDC/CD8+ T-cell MLR assay, we showed that concurrent treatment with DuoBody-CD40×4-1BB and pembrolizumab potentiated IFN-γ, TNF-α and IL-2 production compared with either compound alone (figure 6A, online supplemental figure S5A). This effect was particularly pronounced in the case of IL-2, given its negligible induction by either DuoBody-CD40×4-1BB or pembrolizumab alone. Modulation of targets by DuoBody-CD40×4-1BB treatment in both CD8+ T cells (figure 6B, online supplemental figure S5B) and DCs (online supplemental figure S6) was confirmed by flow cytometry. The observed increase in 4-1BB+ CD8+ T cells and decrease of CD27+ CD8+ T cells induced by DuoBody-CD40×4-1BB were further potentiated by the combination with pembrolizumab (figure 6B, online supplemental figure S5B).

Combination of DuoBody-CD40×4-1BB with pembrolizumab amplifies the magnitude of the immune response. (A,B) Purified CD8+ T cells from healthy donors were cocultured with LPS-matured allogeneic DCs in the presence of DuoBody-CD40×4-1BB, research-grade pembrolizumab (either alone or in combination (concurrent treatment)) or control antibodies for 5 days. (A) Cytokine concentrations in supernatant taken after 5 days of culture in the presence of indicated antibodies. Data shown are mean concentration±SD of duplicate wells from one representative donor pair (donor pair two is shown in online supplemental figure 5). Dashed line indicates cocultures that were not treated with antibody (No Tx). (B) The percentage of 4-1BB+ CD8+ T cells and CD27+ CD8+ T cells was measured by flow cytometry. Data shown are the percentage of positive cells within the total CD8+ T-cell population from one representative donor pair (donor pair two is shown in online supplemental figure 5). Dashed line indicates cocultures on Day 0 (before antibodies were added to the treated cocultures) and cocultures that were not treated with antibody at Day 5 of the MLR assay (No Tx). (C) CD8+ T cells were electroporated with RNA encoding an HLA-A2/CLDN6-specific TCR and PD-1, labeled with CFSE and cocultured with autologous DCs electroporated with CLDN6-encoding RNA in presence of the DuoBody-CD40×4-1BB (0.0022, 0.0067 or 0.2 µg/mL), IgG1 isotype control (0.8 µg/mL), clinical-grade pembrolizumab (0.8 µg/mL) or combinations thereof for 5 days. CFSE dilution in CD8+ T cells was analyzed by flow cytometry. Mean expansion index±SD (n=4) is shown. Dashed line indicates CD8+ T cells that were cocultured with DCs that were not electroporated with CLDN6 in the presence of the IgG1 isotype control antibody as negative control. CFSE, carboxyfluorescein succinimidyl ester; DC, dendritic cell; LPS, lipopolysaccharide; TCR, T-cell receptor.
To further substantiate these findings, we used the CLDN6-specific coculture assay to evaluate T-cell proliferation in vitro. In this setting, DuoBody-CD40×4-1BB or pembrolizumab alone moderately enhanced proliferation (figure 6C). The combination of both agents led to a marked increase in proliferation, even at DuoBody-CD40×4-1BB concentrations that only marginally affected proliferation as a single agent. Together, these data suggest that the combination of DuoBody-CD40×4-1BB with PD-L1/PD-1 blocking antibodies can amplify the magnitude of the immune response.
DuoBody-CD40x4-1BB demonstrated clear peripheral immune modulation in patients with advanced solid tumors
We next evaluated peripheral immune modulation in blood samples from patients with advanced solid tumors that were treated with DuoBody-CD40×4-1BB in the dose-escalation phase of the first-in-human clinical trial (NCT04083599). Combined data from patients treated at dose levels of 30 mg or higher showed that DuoBody-CD40×4-1BB induced a transient increase in peripheral IFNγ and thymus-regulated and activation-regulated chemokine (TARC; CCL17), which peaked on Day 8 after infusion (figure 7A). Furthermore, DuoBody-CD40×4-1BB induced a gradual decrease in absolute peripheral B-cell counts until Cycle 1 Day 8, indicative of B-cell trafficking (figure 7B). This was reciprocated by a gradual increase in the percentage CD86+ B cells in the remaining peripheral B-cell population, indicative of B-cell activation (figure 7B). Finally, a transient increase in the percentage of Ki67+ total and effector memory CD8+ T cells was observed on Cycle 1 Day 8 (figure 7C). All modulated pharmacodynamics returned to baseline levels prior to administration of the second dose of DuoBody-CD40×4-1BB. Collectively, these observations are in agreement with the proposed mechanism of action (MoA), preclinical proof-of-concept results and support the biological activity of DuoBody-CD40×4-1BB in patients with advanced solid tumors.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
DuoBody-CD40×4-1BB mediates broad immune modulation of peripheral APCs and T cells in patients. Patients with advanced solid tumors were treated with DuoBody-CD40×4-1BB in a first-in-human clinical trial (NCT04083599). Data shown are preliminary data from the dose-escalation phase of the ongoing study from patients treated with 30 mg or higher doses of DuoBody-CD40×4-1BB (cut-off date: 20 August 2021). Blood samples were collected on the indicated time points and analyzed for cytokines and immune cell subsets. For each parameter, the mean fold change compared with Cycle 1 Day 1 predose is shown. Numbers in the top right corners denote the number of patients that were included for each parameter. (A) Mean fold change±SEM of IFN-γ and TARC compared with Cycle 1 Day 1 predose. (B) Mean fold change±SEM of absolute CD19+ B-cell counts (cells/μL; calculated using the following formula: (B cells (event count)/leukocytes (event count)*LEUKOCYTES (103 cells/µL))*1000) and the percentage of CD86+ B cells within the CD19+ B-cell population compared with Cycle 1 Day 1 predose. C. Mean fold change±SEM of the percentage of CD8+Ki67+ T cells within the CD8+ T-cell population and CD8+ Ki67+ effector memory T cells within the CD8+CD45RA-CCR7- effector memory T-cell population compared with Cycle 1 Day 1 predose. APC, antigen presenting cell; IFN, interferon.
Discussion
Here, we report preclinical characterization of the MoA of DuoBody-CD40×4-1BB, a novel bsAb developed for the treatment of patients with advanced solid tumors. DuoBody-CD40×4-1BB exhibits conditional CD40 and 4-1BB agonist activity by simultaneous binding to CD40+ APCs and 4-1BB+ T cells. Crosslinking of both targets is essential for CD40 and 4-1BB agonist activity, as FcγR-mediated crosslinking of target-bound antibodies is absent by design. By this unique MoA, DuoBody-CD40×4-1BB induces DC maturation and enhances activation, proliferation and effector functions of human T cells in vitro and promotes TIL expansion ex vivo. In agreement with this preclinical MoA, DuoBody-CD40×4-1BB mediates clear immune modulation of peripheral APCs and T cells in patients with advanced solid tumors. These findings demonstrate that targeting of these two pathways with an Fc-inert bsAb may be an efficacious approach to (re)activate tumor-specific immunity.
The rationale for developing DuoBody-CD40×4-1BB was based on the simultaneous targeting of preclinically validated targets that have shown encouraging clinical activity, despite some limitations. While initial monotherapy response rates of >20% were observed in melanoma patients in clinical trials of CD40 agonistic mAbs, these response rates could not be confirmed in subsequent trials, and response rates were low in nonmelanoma patients with solid tumors.6 Similarly, clinical responses to 4-1BB agonistic mAbs at safe dose levels have been limited, although clinical responses for urelumab at dose levels that were not well-tolerated support the potential benefit of 4-1BB agonists.3 Current clinical strategy for both targets has shifted to exploring various combinations of these agonists with other classes of therapeutic agents, including CPI and chemotherapy. Preclinical evidence suggests that while single agent activity is observed in some models, antitumor activity of CD40 agonists is often dependent on CD8+ T cells.27–29 Furthermore, a 4-1BB-specific monoclonal antibody promotes the generation of tumor-specific immune responses by direct activation of CD8+ T cells in a CD40-dependent manner.30 Therefore, agonist immunotherapies that enhance CD8+ T-cell responses through concomitant CD40 and 4-1BB stimulation may represent a promising strategy to further promote priming and expansion of antitumor immunity.
The FcγR crosslinking-independent bispecific approach presents the advantage that agonist activity is redirected to areas with enhanced expression of both targets, such as tumors and associated lymphoid tissues. Within the TME, 4-1BB is expressed by a CD8+ T-cell subset that is enriched for tumor antigen-specific cells.31–33 CD40 expression has been reported in many solid tumor indications, either on tumor-infiltrating DCs or on cancer cells,34–39 which can provide the required crosslinking for DuoBody-CD40×4-1BB. In addition to intratumoral activity, we hypothesize that the tumor-draining lymph node is an important site of action for DuoBody-CD40×4-1BB to enhance T-cell activation, due to the presentation of tumor antigens by CD40+ APCs to 4-1BB+ T cells. Our findings support this hypothesis as conditional 4-1BB agonist activity critically depends on previous TCR stimulation. Therefore, only recently primed T cells can be targeted by this means.
Targeting DCs has attracted high interest for cancer immunotherapy, owing to their central role in orchestrating anti-tumor immunity by priming tumor-specific T cells in tumor-associated LNs.40 DC-targeting agents that are currently in clinical use include Toll-like receptor agonists, tumor-associated antigen vaccines and immunostimulatory cytokines. These agents primarily act by mobilizing and activating DCs, which subsequently enhances tumor-specific T-cell responses.40 CD40 agonists improve tumor-specific T-cell immunity independently of innate immune receptors by inducing DC maturation and secretion of proinflammatory cytokines.27 41 42 Accordingly, antitumor activity of CD40 agonist immunotherapy critically depends on cross-presenting DCs and CD8+ T cells in preclinical models.28 29 41 Indeed, we show that DuoBody-CD40×4-1BB enhanced DC maturation through crosslinking with CD8+ T cells in the absence of exogenous innate immune stimuli. The use of a bispecific antibody could be advantageous in this context, as it directs CD40 agonist activity to areas of T-cell activation, that is, areas with activation-induced 4-1BB expression, thereby providing a more focused stimulus for DC maturation. Hence, DuoBody-CD40×4-1BB-mediated DC crosslinking with 4-1BB+ CD8+ T cells may provide a substantial benefit for the priming and amplification of tumor-specific CD8+ T-cell responses.
Bispecific antibodies with conditional 4-1BB agonistic activity currently in preclinical and clinical development include agents that target agonist activity to intratumoral expressed proteins (eg, Her2, EGFR and FAP)12 13 15 or that simultaneously block tumor-associated inhibitory pathways (eg, PD-L1).14 Conditional 4-1BB agonist activity is hypothesized to facilitate antitumor activity at safe dose levels, thereby broadening the therapeutic window compared with classical 4-1BB agonistic mAbs. Combining 4-1BB agonist activity with other TNFRs may provide additional co-stimulation and further boost antitumor immunity, as shown preclinically for an Fc-inert CD137xOX40 bispecific antibody.43 CD40 agonism has also been paired with the tumor-specific antigen mesothelin in a bispecific format, which activated T-cell responses via conditional stimulation of CD40+ APCs.44 The CD40xmesothelin bispecific antibody exhibited antitumor activity against mesothelin-expressing tumors in preclinical models and an acceptable clinical safety profile in a Phase I trial.44 45 Similarly, a FAPxCD40 bispecific antibody induced a long-lasting antitumor immune response and exhibited potent antitumor activity in high FAP-expressing tumors, with no signs of toxicity in preclinical models.46 47 While the combination of CD40 and 4-1BB mAbs has not been evaluated clinically, the underlying biology of these two pathways presents with the opportunity to simultaneously enhance T-cell and DC function and provides a clear rationale for combination approaches. Indeed, in a preclinical model, the combination of agonistic mAbs targeting CD40 and 4-1BB resulted in a fourfold enhancement of a vaccine-induced CD8+ T‐cell response compared with either mAb alone.48 We now demonstrate that DC and T-cell functions can be enhanced by a bispecific antibody that exhibits agonist activity by crosslinking CD40 and 4-1BB. These preclinical findings provide in vitro proof-of-concept for the bispecific format confirmed with the initial results from the dose-escalation phase of the first-in-human trial in patients with advanced solid tumors. At biologically active dose levels, DuoBody-CD40×4-1BB induced peripheral T-cell and APC/DC-associated cytokines, trafficking and activation of B cells, as well as increased proliferation of CD8+ T cells. Ongoing analyses in this and subsequent studies will investigate associations with anti-tumor activity.
The bispecific format also offers the potential for further combinations with CPI, chemotherapy or vaccination approaches. As tumor antigen-specific T cells express both PD-1 and 4-1BB,31 33 blockade of PD-L1/PD-1 signals and conditional 4-1BB agonist activity may synergize to enhance T-cell effector functions and antitumor immunity. Here, we provide rationale for combining DuoBody-CD40×4-1BB with PD-1 blocking antibodies, as this combination potentiates multiple biological features to amplify the immune response in vitro. The combination of DuoBody-CD40×4-1BB with pembrolizumab enhanced production of proinflammatory cytokines and T-cell proliferation compared with each monotherapy alone, indicative of a general increase in T-cell activation. The observed increase in 4-1BB and loss of CD27 expression on CD8+ T cells in the MLR assay further suggests that the combination of DuoBody-CD40×4-1BB with pembrolizumab may enhance the shift from naïve CD8+ T cells towards effector (memory) CD8+ T cells,3 49 although this requires a more in depth analysis of cell surface markers after prolonged culture. This improved modulation may allow for enhanced anti-tumor immunity in malignancies that are early stage or resistant/refractory to CPI treatment.
In summary, we have shown that DuoBody-CD40×4-1BB activates the immune system through conditional CD40 and 4-1BB agonist activity. DuoBody-CD40×4-1BB is being jointly developed by Genmab and BioNTech for the treatment of solid tumors. A first-in-human clinical trial to evaluate the safety and biological activity of DuoBody-CD40×4-1BB in patients with advanced solid tumors is currently ongoing (NCT04083599).
Data availability statement
Data are available on reasonable request. The datasets generated during and/or analyzed during the current study are available from the corresponding author on reasonable request.
Ethics statements
Patient consent for publication
Ethics approval
The ongoing first-in-human clinical study (registered on https://www.clinicaltrials.gov by identifier NCT04083599) was approved by appropriate Institutional Review Boards at each participating site and conducted in compliance with the principles of the Declaration of Helsinki, International Council for Harmonisation GCP (ICH GCP E6[R2]) and applicable regulatory requirements. All patients provided written informed consent. Participants gave informed consent to participate in the study before taking part.
Acknowledgments
The authors would like to acknowledge Steven Zenner for his contribution to the imaging studies, Brandon Higgs and Jaime Bald for their assistance on analyzing clinical samples, and Ann-Kathrin Wallisch, Julia Mühl, Maren Köhne, Patrick Goldberg, Christopher Chiu, Rachelle Kosoff, René Völker-van der Meijden, Grietje Andringa, Lucia Rodriguez Rodriguez, Rajaa Snijdewint-Nkairi and Petra Adams-Quack for technical expertise and support.
References
Supplementary materials
Supplementary Data
This web only file has been produced by the BMJ Publishing Group from an electronic file supplied by the author(s) and has not been edited for content.
Footnotes
Correction notice This article has been updated since it was first published online. The author Homer C Adams III has been updated to Homer C Adams 3rd.
Contributors AM, HCA III, FG, KS, IA, JMB, MF, MD, SK, TA, ECWB, DPES, ÖT, AKS, US, MJ-K contributed to conception or design of the work. AM, IA, FG, KS, BS, JQ, MD, SMB, AT, SK, DV, JMB, VMS, MF, ES, FV, HCA III, ECWB and MJ-K contributed to acquisition, analysis or interpretation of the data. SMB and AT have drafted the work and all of the authors have revised the draft. MJ- K acts as the guarantor of the study.
Funding This study was funded by Genmab and BioNTech.
Competing interests US and ÖT are management board members and employees at BioNTech (Mainz, Germany). AM, FG, KS, AT, BS, JQ, MD and SK are employees at BioNTech. Some of the authors have securities from BioNTech. ES and FV are employees at TRON. HCA III, IA, JMB, SMB, DV, VMS, MF, DPES, SK, TA, ECWB and MJ-K are employees at Genmab and own stock and/or stock options. HCA III, IA, DPES, US, AM and FG are inventors on patents and patent applications related to CD40×4-1BB bispecific antibodies.
Provenance and peer review Not commissioned; externally peer reviewed.
Author note AM and HCA III are joint first authors. US and MJ-K are joint last authors.
Supplemental material This content has been supplied by the author(s). It has not been vetted by BMJ Publishing Group Limited (BMJ) and may not have been peer-reviewed. Any opinions or recommendations discussed are solely those of the author(s) and are not endorsed by BMJ. BMJ disclaims all liability and responsibility arising from any reliance placed on the content. Where the content includes any translated material, BMJ does not warrant the accuracy and reliability of the translations (including but not limited to local regulations, clinical guidelines, terminology, drug names and drug dosages), and is not responsible for any error and/or omissions arising from translation and adaptation or otherwise.