Article Text

Statistics from Altmetric.com
- Green Fluorescence Protein
- Tumor Microenvironment
- Intratumoral Administration
- Mouse Glioma
- Glioma Microenvironment
Meeting abstracts
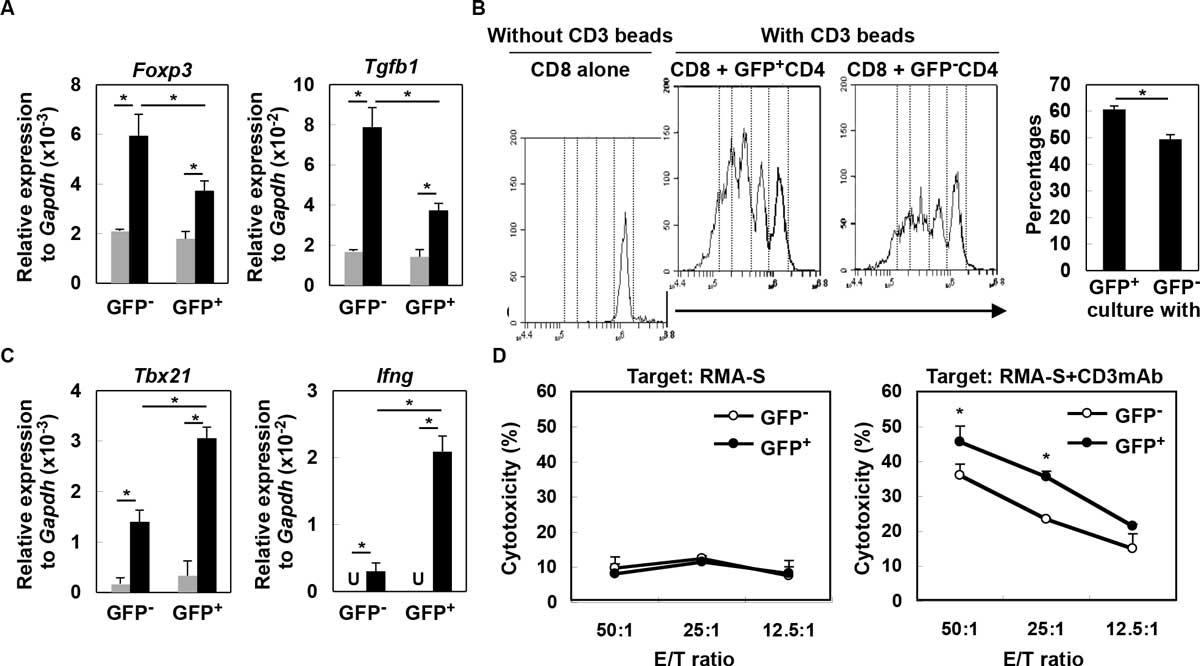
While type-I interferons (IFNs) play critical roles in antiviral and antitumor activity, it remains to be elucidated how type-I IFNs are produced in sterile conditions of the tumor microenvironment and directly impacts tumor-infiltrating immune cells. We report that both human and de novo mouse gliomas show increased expression of type-I IFN messages, and in mice, CD11b+ brain-infiltrating leukocytes (BILs) are the main source of type-I IFNs that is induced partially in a STING (stimulator of IFN genes)-dependent manner. Consequently, glioma-bearing StingGt/Gt mice showed shorter survival, and lower expression levels of Ifns compared with wild-type mice. Furthermore, BILs of StingGt/Gt mice show increased CD11b+ Gr-1+ immature myeloid suppressor and CD25+ Foxp3+ regulatory T (Treg) cells, while decreased IFN-γ-producing CD8+ T cells. To determine the effects of type-I IFN expression in the glioma microenvironment, we utilized a novel reporter mouse model, in which the type-I IFN signaling induces the Mx1 (IFN-induced GTP-binding protein) promoter-driven Cre recombinase, which turns the expression of loxp-flanked tdTomato off, and turns green fluorescence protein (GFP) expression on, thereby enabling us to monitor the induction and effects of IFN signaling in the glioma microenvironment. CD4+ T cells that received direct type-I IFN signals (i.e., GFP+ cells) demonstrate lesser degrees of regulatory activity based on lower Foxp3 and Tgfb1 expression levels (Figure 1) as well as lesser suppression of CD8+ T cell proliferation (Figure B). IFN-sensed CD8+ T cells exhibit enhanced levels of Th1 markers, Tbx21 and Igfng (Figure C), as well as cytotoxic T-cell activity based on reverse antibody-dependent T-cell-mediated cytotoxicity assay (Figure D). Finally, intratumoral administration of a STING agonist (cyclic diguanylate monophosphate; c-di-GMP) improves the survival of glioma-bearing mice associated with enhanced type-I IFN signaling, Cxcl10 and Ccl5 and T cell migration into the brain. In a combination with subcutaneous OVA peptide-vaccination, c-di-GMP increased OVA-specific cytotoxicity of BILs and prolonged the survival. These data demonstrate significant contributions of STING to antitumor immunity via enhancement of the type-I IFN signaling in the tumor microenvironment, and imply a potential use of STING agonists for development of effective immunotherapy, such as the combination with antigen-specific vaccinations.

{kind=link}
Type-I IFNs directly impact on T-cell functions in glioma-developing mice. (A) CD4+ cells from draining LN derived from glioma-developing tdTomato mice were sorted into GFP- or GFP+ cells and incubated with (black bars) or without (grey bars) anti-CD3mAb. After 4 h, total RNA was extracted for evaluation of Foxp3 and Tgfb1 mRNA levels by qRT-PCR. (B) CFSE-labeled WT CD8+ T-cells were co-cultured with GFP- or GFP+ CD4+ T-cells in the presence of CD3 beads. After 60 h, division of CFSE-labeled CD8+ T-cells gated by reactivity to PE-Cy7-condjugated anti-CD8mAb was evaluated by CFSE intensity. As a negative control, CFSE-labeled WT CD8+ T-cells were cultured without any stimulation (left panel). Histograms are representative of two independent experiments. The bar graph shows the percentage of CD8+ cells that have divided at least twice in each of two stimulation conditions (N = 4/group; *p < 0.05). (C) GFP- or GFP+ CD8+ T-cells were incubated with (black bar) or without (grey bar) anti-CD3mAb. After 4 h, total RNA was extracted for evaluation of Tbx21 and Ifng mRNA expression levels by qRT-PCR (U: undetected). (D) Cytotoxic activity of GFP- and GFP+ CD8+ T-cells was evaluated by 51Cr-release assay. RMA-S cells untreated (left panel) or pretreated (right panel) with anti-CD3mAb (10 g/mL) were used as target cells. *p < 0.05 compared at the same E/T ratio.