Article Text

Abstract
Objectives Emerging evidence has shown a role for tumor antigen-specific regulation in cancer. Identifying individuals with pre-existing regulatory responses may be key to understand those who are more likely to respond to Programmed Death-1 (PD-1) or PD-1 Ligand 1 (PD-L1) checkpoint blockade. We hypothesized that a functional assay could identify the role of PD-1/PD-L1 interactions on tumor-specific immune cells in the peripheral blood in patients with advanced non-small-cell lung cancer (NSCLC).
Methods We performed the trans vivo delayed-type hypersensitivity assay to identify the role of PD-1/PD-L1-mediated tumor-specific immune regulation in ten patients with advanced NSCLC.
Results The majority of patients had PD-1-mediated anergic immune responses towards their tumor antigens. Eight out of nine of these patients did not respond to their own tumor antigens but responded in the presence of anti-PD-1 antibody (‘PD-1 anergy’ phenotype). A minority (3/9) also had ‘active’ PD-1-mediated immune suppressive regulatory responses. Our results suggest that PD-1-anergy is a common feature of NSCLC immune responses, whereas PD-1-mediated immune suppression is present only in a minority of patients. The latter was associated with poor clinical outcomes in our sample.
Conclusions Overall, our results indicate that bystander suppression or the ‘anergy-only’ phenomenon may be novel biomarkers in NSCLC and suggest prediction value based on these phenotypes.
- immunology
- biomarkers, tumor
- costimulatory and inhibitory T-cell receptors
- programmed cell death 1 receptor
- lung neoplasms
This is an open access article distributed in accordance with the Creative Commons Attribution Non Commercial (CC BY-NC 4.0) license, which permits others to distribute, remix, adapt, build upon this work non-commercially, and license their derivative works on different terms, provided the original work is properly cited, appropriate credit is given, any changes made indicated, and the use is non-commercial. See http://creativecommons.org/licenses/by-nc/4.0/.
Statistics from Altmetric.com
- immunology
- biomarkers, tumor
- costimulatory and inhibitory T-cell receptors
- programmed cell death 1 receptor
- lung neoplasms
Introduction
Immune checkpoint inhibitor (ICPI) therapy has revolutionized non-small-cell lung cancer (NSCLC) treatment and is now standard in advanced NSCLC. Several pivotal studies have demonstrated the survival benefit of ICPIs, including KEYNOTE-024,1 KEYNOTE-189,2 IMpower-150,3 and KEYNOTE-407.4 KEYNOTE-042 has led to approval for first-line pembrolizumab in a broader group of patients with 1% or greater Programmed Death Ligand 1 (PD-L1 expression).5 A Global Study to Assess the Effects of MEDI4736 Following Concurrent Chemoradiation in Patients With Stage III Unresectable Non-Small Cell Lung Cancer (the PACIFIC trial) established a role for earlier use of checkpoint inhibition, demonstrating improved overall survival with durvalumab after chemoradiation in stage III NSCLC.6
A major challenge remains identifying patients most likely to benefit from ICPI. In the aforementioned studies, higher PD-L1 expression correlates with improved responses to PD-1/PD-L1 pathway blockade; however, it is not the sole predictor of response. For example, microsatellite instability,7 tumor mutational burden, and a T-cell inflamed gene signature can predict responses to ICPI.3 8–10 Furthermore, the expression of PD-L1 can display spatiotemporal variability,10 11 and in consequence, one biopsy may not represent global tumor expression. As such, PD-L1 has limitations as a predictive biomarker, and therefore a functional immune assay may accurately measure the immunological activity of individual patients. As patients develop ICPI refractory disease, dynamic biomarkers may inform subsequent therapy.
A more complete understanding of the role of PD-1/PD-L1 interactions in the tumor microenvironment may improve patient selection for checkpoint blockade. The inconsistent correlation between tumor PD-L1 expression and response to checkpoint inhibition underlines the complexity of regulation of the antitumor immune response by the PD-1 pathway and the consequences of relief of this suppression. One potential explanation is the cell population that expresses PD-L1, as both tumor cell and tumor-infiltrating leukocyte (TIL)-associated PD-L1 have critical roles in tumor immune evasion.12 13 It is likely that coinhibitory molecules will be more effective mediators of antitumor immune suppression when expressed both in effector and regulatory cells, as T-regulatory cells mediate ‘active’ immune suppression rather than ‘passive’ anergy of effector T cells alone.
The trans vivo delayed-type hypersensitivity (tvDTH) assay has been used to characterize regulation systems in transplant recipients14 15 and in prostate cancer.16 Delayed-type hypersensitivity (DTH) responses are a standard method to evaluate cellular immune responses in animal and human immunotherapy trials.17 Thus, we hypothesized that the tvDTH will accurately describe the contribution of PD-1/PD-L1 interactions in regulating immune responses towards tumor antigens. This functional assay directly shows modifications of tumor-specific immune responses induced by checkpoint blockade and may augment PD-L1 immunohistochemistry as a biomarker.18
Materials and methods
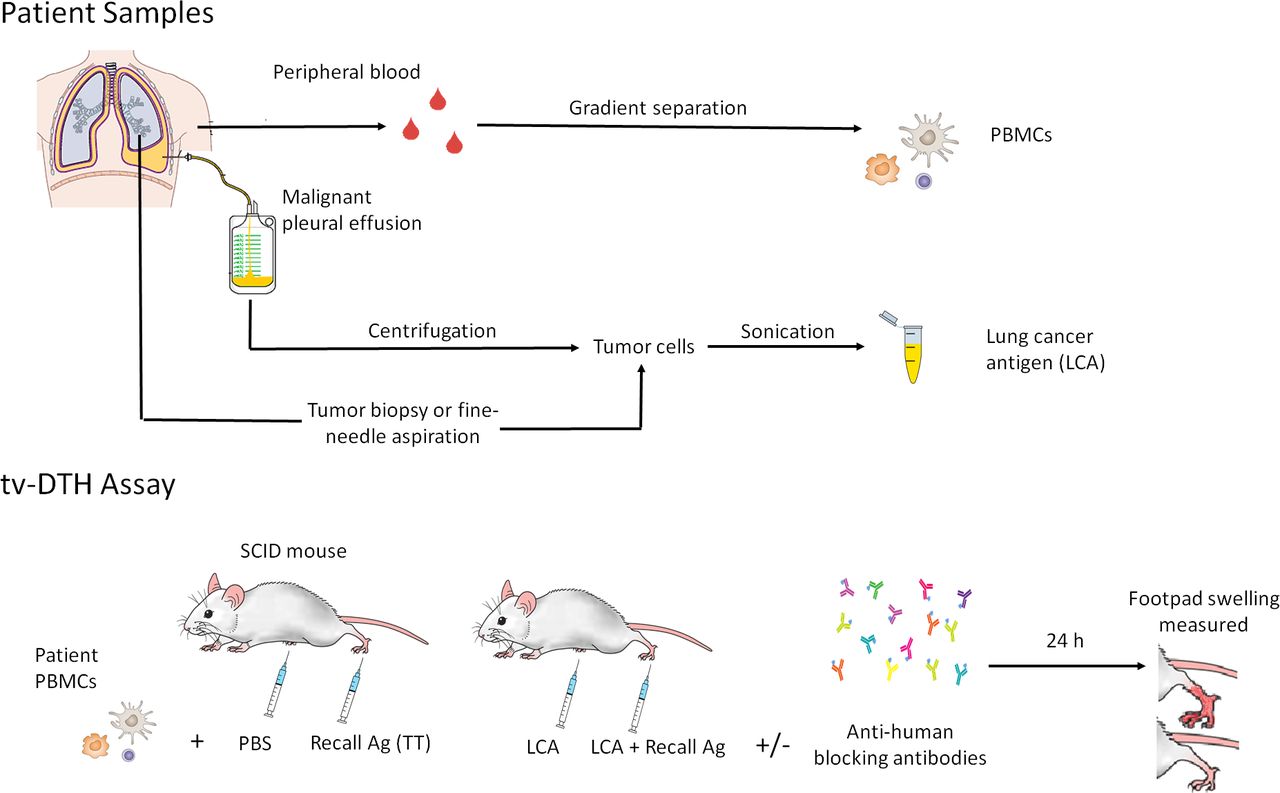
In tvDTH assays, the components of a DTH (immune cells and antigen) are transferred to footpads of immunodeficient mice. Malignant cells from fine-needle aspirates or pleural effusions (centrifuged at 2000 g for 10 min to pellet the cells in suspension) were sonicated as the source of tumor antigen (Ag) preparations (figure 1A). The presence of malignant cells was confirmed by a pathologist.

Workflow of patient sample processing. (A) Patients are immunized with TT/DT 2 weeks prior to obtaining a blood draw. LCA is obtained from malignant pleural effusion cell pellet or fresh tumor sample (B) tvDTH assay summary. SCID mice footpads are injected with patient PBMCs+tumor Ag and footpad swelling measured after 24 hours. A replicate set of footpad conditions, including antihuman PD-1 (pembrolizumab) and/or CTLA-4 (AS32) blocking antibodies, is performed to study the role of these molecules. CTLA-4, cytotoxic T lymphocyte associated protein 4; LCA, lung cancer antigen; PBS, Phosphate buffered saline;PBMC, peripheral blood mononuclear cell; SCID, severe combined immunodeficient; TT/DT, tetanus/diphtheria; tvDTH, trans vivo delayed-type hypersensitivity.
Peripheral blood was collected from nine patients with NSCLC in conjunction with routine clinical labs at the University of Wisconsin Carbone Cancer Center. Peripheral blood mononuclear cells (PBMCs) were isolated by Ficoll gradient separation. Patients underwent a tetanus/diphtheria (TT/DT) vaccination 2 weeks prior to blood draws, which served as a positive control recall antigen.
PBMCs were challenged with specific antigens, inducing an inflammatory cascade in mouse footpads, which can be measured as a swelling response. This response is antigen-specific and requires prior antigen sensitization. The role of specific molecules can be interrogated by coinjecting blocking antibodies. Thus, this assay is suitable to investigate mechanisms controlling effector and regulatory antigen-specific immune responses (figure 1B).
Seven million PBMCs were injected into footpads of 6–8 week CB.17-SCID mice (Prkdcscid lymphopenic, hypogammaglobulinemic mice lacking functional T and B cells), together with 10 µg of tumor Ag preparation (derived from patients’ own tumors). PBMCs plus phosphate-buffered saline (PBS) was used as a negative control, and response to TT/DT (Aventis Pasteur, Bridgewater, New Jersey, USA) plus PBMCs was used as a positive control. DTH reactivity was measured after 24 hours as the change in footpad thickness using a dial thickness gage (Mitutoyo, Kawasaki, Japan). Net swelling was determined by subtracting ‘background’ swelling of a control injection of PBMC plus PBS.
To investigate the role of immunoregulatory receptors on these responses, 1 µg of humanized anti-human PD-1 (Keytruda (pembrolizumab), Merck) and/or murine antihuman CTLA-4 (clone AS32; Ab Solutions, Mountain View, California, USA) were coinjected in a replicate set of footpad conditions to block these receptors and to study their contribution to the swelling response. Anergy was operationally defined as an increase in Ag-induced footpad swelling when a checkpoint is blocked. Detailed tvDTH methodology has been described previously.19
We determined the bystander inhibition of recall responses to TT/DT in the presence of tumor antigens by comparing the net swelling of each injection using the following formula:

Bystander suppression was operationally defined as a reduction of TT/DT-induced swelling when Ag was coinjected.
Statistical analysis was performed using GraphPad Prism V.6.05. Unpaired t-tests were used to compare DTH swelling responses between patients, while paired t-tests were used to compare DTH swelling differences for each patient under different footpad conditions.
Results
Nine patients with advanced NSCLC (stage III/IV) from February 2017 to December 2018 enrolled and consented to the study. Patient characteristics, demographics, stage, tumor histology and PD-L1 expression status are shown in table 1. The median age was 65 years; 55% were female; and 88% had a smoking history. The majority of patients had adenocarcinoma histology (77%).
Summary of clinical characteristics and trans vivo delayed-type hypersensitivity responses of the patient cohort
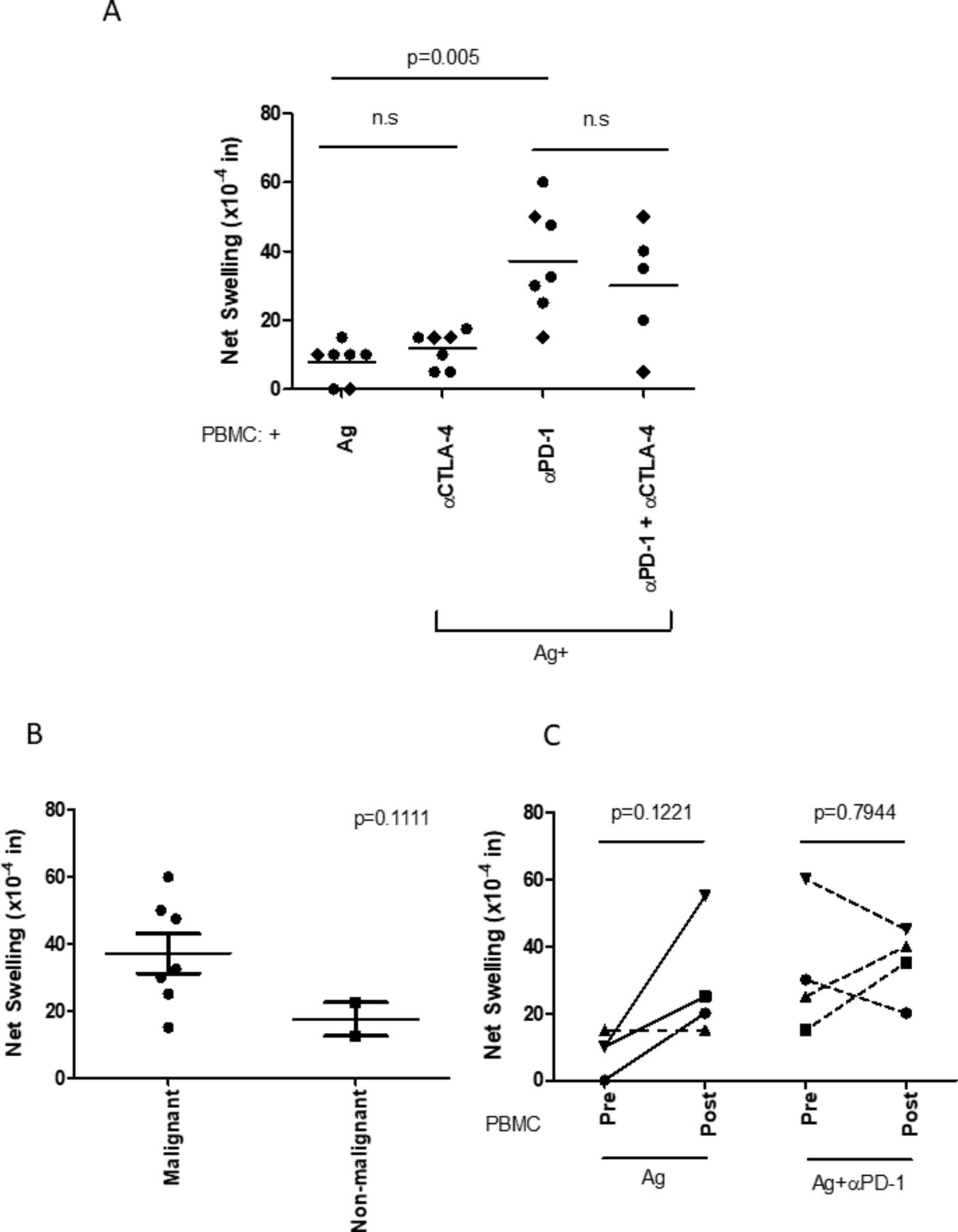
We observed little to no antigen-specific swelling (7.85×10–4 inches, SD=±5.66; figure 2A) when PBMCs were challenged with tumor Ag alone. This could indicate that tumor Ag is weakly immunogenic or that immune regulatory mechanisms exist in these patients, which suppress immune responses towards tumor Ag.

Tumor antigen-specific effector immune responses are inhibited by the PD-1 pathway in patients with NSCLC. (A) PBMCs from patients with NSCLC challenged with antigen prepared from their own tumor cells (with malignant cells confirmed by pathology, n=5–7) produced minimal DTH swelling. However, PD-1 blockade uncovered potent tumor-specific responses. CTLA-4 blockade alone was less effective at doing so either by itself or when coinjected with αPD-1 antibody. (B) PBMCs from patients with NSCLC were challenged with antigen preparations derived from samples with reported presence or absence of malignant cells by pathology, plus anti-PD-1 antibody. PD-1 blockade uncovered potent antigen-specific responses with malignant samples, which were in most cases stronger than those with non-malignant samples. (C) Pretreatment versus post-treatment tumor antigen-induced swelling. For four patients, we obtained blood samples before and after treatment. We observed that, in all cases but one, the post-treatment PBMCs induced greater DTH swelling responses when challenged with tumor antigen. However, the antigen plus anti-PD-1-induced swelling was similar at both timepoints. ●, adenocarcinoma; ♦, squamous cell carcinoma; ns, not significant; NSCLC, non-small-cell lung cancer; PBMC, peripheral blood mononuclear cell.
We were interested in studying the role of PD-1/PD-L1 and CTLA-4 as potential mediators of this phenomenon due to their therapeutic relevance in NSCLC. To study inhibitory cosignaling molecules in regulating the NSCLC antigen-specific immune response, 1 µg of antihuman PD-1 (pembrolizumab) or antihuman CTLA-4 (AS32) was coinjected in a replicate set of footpad conditions. We observed that, in all cases but one, blocking PD-1 resulted in potent swelling responses (37.14×10–4 inches, SD +±15.84), which was statistically significant (p=0.005) (figure 2A). In contrast, CTLA-4 blockade did not significantly increase the swelling response (p=0.157) against tumor Ag, either by itself or when coinjected with anti-PD-1 (αPD-1) antibody (p=0.2992).
Importantly, as an autoantigen control, we performed the tvDTH assay using antigen preparations derived from patient pleural effusion samples with no malignant cells identified by cytopathology. In most cases, antigen preparations from samples positive for malignant cells induced greater DTH swelling (figure 2B). Tumor PD-L1 expression on immunohistochemistry bore no relationship to PD-1-mediated tvDTH swelling (data not shown). Thus, we concluded that NSCLC tumor Ag are immunogenic and commonly regulated through PD-1 in this cohort of patients.
We then asked if treatment modified antitumor responses in patients with NSCLC. We had four patients in whom we repeated a tvDTH assay after initiating standard or experimental treatment. We found that tumor antigen-specific DTH responses in three out of four patients increased, with an average of 20×10–4 inches (p=0.12) from the pretreatment to the post-treatment timepoint (figure 2C). The only exception was patient 10 (dashed line), who did not show any PD1-mediated anergy at the initial time point. This patient had rapid progression of disease during first-line anti-PD1 therapy. In contrast, no consistent trends were identified in the PD-1-blocked swelling before and after treatment (figure 2C). This could be due to the heterogeneity of this patient subset, with two patients receiving PD-1 checkpoint inhibitors, one receiving an ALK inhibitor and one undergoing chemoradiation.
Having found that PD-1 interactions inhibited immune responses to autologous tumor Ag in nearly all of the patients with NSCLC prior to treatment, and that therapy restored tumor Ag-directed responses in those patients who previously showed PD1-dependent inhibition, we inquired whether these effects were due to ICPI impact on effector T cells, regulatory cells, or both. The first possibility—that PD-1 interactions on effector cells alone suppressed their function—would be indicative of anergy (ie, state of immune unresponsiveness induced in T cells associated with increased expression of immunoregulatory receptors and dysfunction). Alternatively, the increase in DTH responses observed after PD-1 blockade could be due to targeting PD-1 on regulatory cells. These possibilities differ in that, in anergy, PD-1 expression occurs in the effector cell and directly inhibits it, while in suppressive responses, PD-1 expression on regulatory cells is necessary for active immune suppressive mechanisms to inhibit another (effector) cell. Thus, these regulatory cells could suppress bystander effector responses. A third possibility is that both anergy and immune suppression are happening simultaneously.
To investigate this, we coinjected in the mouse footpads TT/DT (recall antigens to which the patients were immunized) either alone or together with tumor Ag.
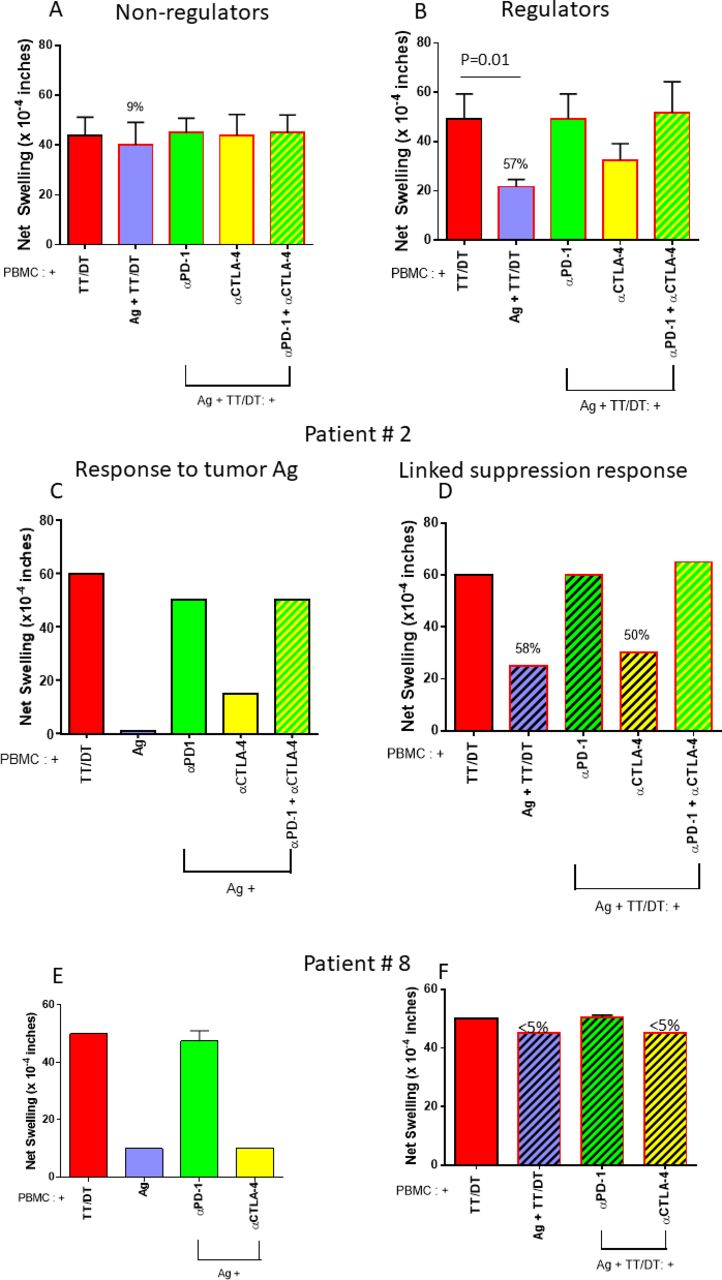
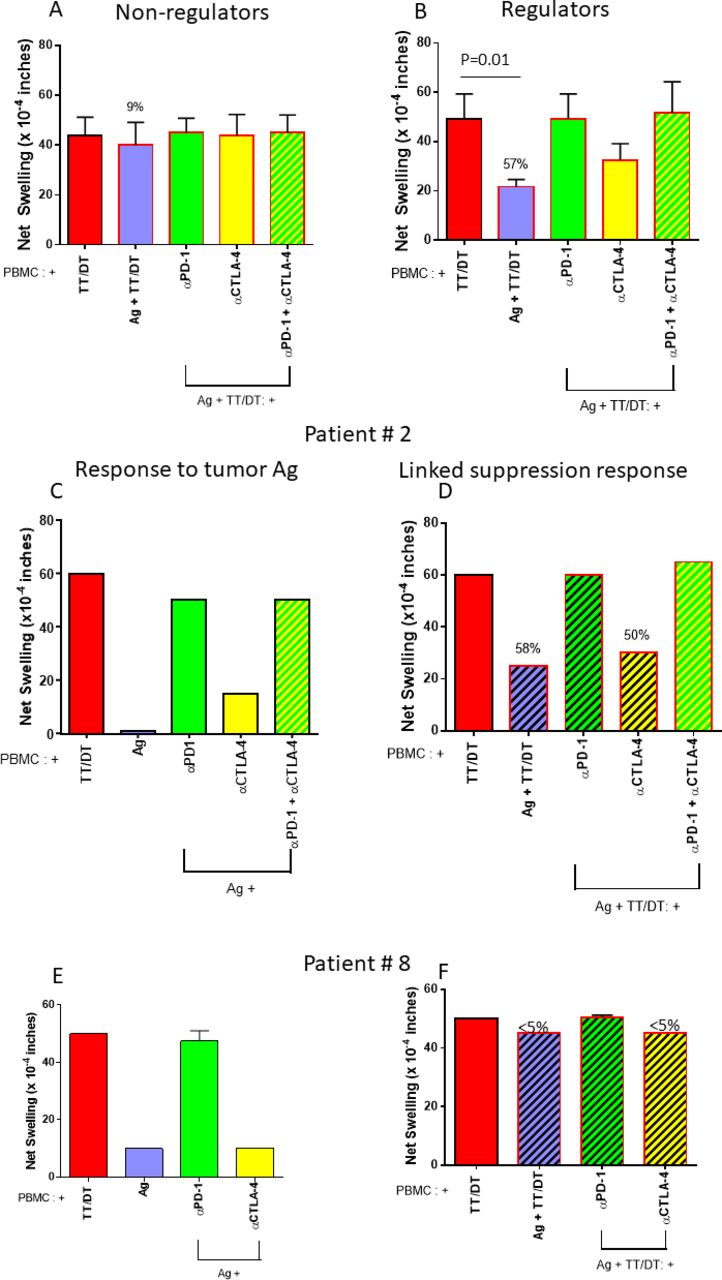
TT/DT by itself elicited robust swelling responses. In three out of nine patients, however, we observed that coinjecting TT/DT plus tumor Ag reduced the swelling response, as compared with TT/DT by itself (figure 3A,B). PBMC from these ‘regulator’ patients reduced the TT/DT-specific swelling response significantly more than ‘non-regulators’, for which tumor Ag did not modify the TT/DT-induced response (compare figure 3A,B; p=0.01). This indicated the presence of tumor antigen-specific regulatory cells which are able to actively suppress bystander effector immune responses. Further, this reduction in swelling response to TT/DT was abrogated when PD-1 (but not CTLA-4) was blocked, indicating that these regulatory cells use PD-1. In these patients, PD-1-dependent regulation could have contributed to the uncovering of antigen-specific swelling when PD-1 was blocked. Hence, we detected PD-1-dependent suppressive responses towards tumor Ag in three out of nine patients with NSCLC in our cohort.
{kind=link}
{kind=link}
{kind=link}
Bystander regulation is present in a subset of patients. (A) PBMCs obtained from six patients with NSCLC showed robust TT/DT-induced DTH swelling responses that were not significantly modified by tumor antigen or checkpoint blockade. (B) In three cases, however, coinjection of tumor antigen dramatically reduced TT/DT-induced swelling, which returned to baseline after PD-1, but not CTLA-4, blockade. This indicates the existence of PD-1-mediated tumor antigen-specific regulatory responses in a minority of patients with NSCLC. DTH profile examples: examples of DTH profiles from two patients. (C,D) Profile from patient 2. C depicts response to antigen alone or with anti-PD-1, anti-CTLA4 inhibitors, or both. This patient showed minimal response towards tumor antigens. However, PD-1 blockade uncovered a potent swelling response, while CTLA-4 blockade had a more modest effect. (D) Response to antigen and TT together is depicted, and in this patient, coinjection of tumor antigen reduced the TT/DT response by 58%. Anti-PD-1 antibody, but not anti-CTLA4 antibody, abrogated the tumor antigen induced bystander suppression. Altogether, these data are consistent with PD-1-dependent anergy and linked suppression towards tumor antigens. (E,F) Profile from patient 8 DTH profile. (E) A modest tumor antigen-specific response was observed that was greatly enhanced by blocking PD-1, but not CTLA-4. (F) No bystander suppression of recall antigen-induced swelling was present. This depicts a typical DTH pattern of PD-1-mediated anergy towards tumor antigens, without bystander suppression. PBMC, peripheral blood mononuclear cell; TT/DT, tetanus/diphtheria.
Figure 3C–F shows representative tvDTH response patterns in two patients. Patient 2 showed bystander suppression as indicated by Ag-mediated reduction of TT/DT-induced swelling, which was reversed with anti-PD1 treatment. This patient had progressive intracranial disease with rapid functional decline that precluded next line therapy with ICPI. In contrast, patient 8 did not display bystander suppression, and the Ag-mediated suppression was reversed with anti-PD1 therapy. This patient completed concurrent chemoradiation and consolidative durvalumab and remains on surveillance, with no definite evidence of disease progression.
Discussion
Analysis of our cohort of nine patients with advanced NSCLC allowed us to draw several conclusions. First, we detected PD-1-mediated anergy towards tumor Ag in eight out of nine patients. This suggests that the presence of tumor-specific, anergic T cells expressing PD-1 may be a common feature of NSCLC. In contrast, CTLA-4-mediated anergy was not observed. Second, a subset of patients further demonstrated PD-1-dependent bystander suppression. This suggests the existence of regulatory cells that use the PD-1/PD-L1 axis to suppress tumor antigen-specific effector T cells. Currently, we do not know the phenotypes of these cells, which might include regulatory T cells, or possibly another population. In a previous study where we performed tvDTH on prostate cancer patients, we determined that the suppressive cell population was a CTLA-4+, interleukin-35-producing CD8 T cell in that setting.16 We observed the regulator pattern in three patients in our cohort. Notably, all of these patients had poor clinical outcomes. Altogether, our preliminary results are consistent with a hypothesis that bystander suppression serves as an additional layer of immune regulation. This implies more effective tumor immune escape in the context of immune regulation plus anergy, potentially leading to poorer clinical outcomes as compared with anergy alone. However, these findings need additional validation in a larger prospective cohort.
It is interesting that in three out of four patients, a repeat tvDTH after they had initiated treatment showed an increase in the antigen-specific swelling response. This could reflect relief of malignancy-associated immune suppression by antineoplastic treatment not limited to immune therapy, because this subset included patients treated with checkpoint inhibitors, chemoradiation, or an ALK inhibitor. The patient who did not show such augmentation had rapid disease progression. The change in tvDTH responses before and after treatment may reflect functional changes in patients’ immune activation status and underscores the dynamic nature of the assay.
Ultimately, the tumor immune response is orchestrated by TILs; however, obtaining sufficient TILs from patients is difficult to do routinely. Therefore, we hypothesized that interrogating tumor antigen-specific circulating immune cells could be useful to probe the tumor immune response using a non-invasive procedure (phlebotomy). Future studies comparing tvDTH profiles using patient-matched TILs and PBMCs could shed light into differences between these lymphocyte subsets. There have been efforts to identify phenotypes of circulating cells relevant to the tumor immune response that are predictive biomarkers of immunotherapy response in NSCLC. Kamphorst and colleagues observed that increases in proliferating peripheral PD-1 +CD8 T cells after PD-1 blockade predicted positive responses.20 We have identified prostate cancer immune escape mechanisms after prostatic acid phosphatase (PAP) vaccination by performing DTH using circulating immune cells.16 Here, we identified modulation of tvDTH responses of tumor antigen-specific immune cells in NSCLC PBMCs by PD-1. The presence of PD-1-mediated anergy alone, without PD-1-mediated immune suppression, appeared to predict a successful outcome of therapy. It also suggests that the presence of additional depth of immune regulation in NSCLC indicates more effective tumor immune escape that is harder to overcome and predicts poor prognosis. Our results suggest that bystander immune suppression may be an informative additional biomarker. Further investigation with a larger study cohort is needed to ascertain whether it can predict response to ICPI or if it indicates a poor prognosis in general.
There are limitations to our study. Our exploratory investigation enrolled a small cohort of heterogeneous patients with advanced NSCLC. Nonetheless, it seems remarkable that all but one patient exhibited an antitumor response when disinhibited by PD-1 blockade at their initial recruitment time point. Another limitation relates to standardization of tumor antigens to challenge immune cells and the number of cells recovered from each patient. In this study, tumor Ag was prepared from confirmed malignant cells, and equal amounts of protein used to stimulate patient PBMCs, and as such, it is difficult to determine and standardize the neoepitope load in our tumor lysates. This is inherent to the heterogeneity of neopitopes in between patients with NSCLC, with limited common antigens specific for NSCLC.21 However, patient-derived tumor Ag preparations could stimulate antigen-specific cells unique to each patient, and this approach has been used to examine memory T-cell function in breast and pancreatic cancer.22 23 The lesser uncovered response in the presence of anti-PD1 antibody in the case of two patients tested with self antigen prepared from a non-tumor sample (figure 2B) argues that the uncovered responses were ‘tumor-specific’ for each patient.
This pilot study is the first report of use of the tvDTH assay to evaluate immune activation status in patients with NSCLC. Results demonstrate that the tvDTH could be used to identify mechanisms of immune regulation and exhaustion. Our initial data raise the hypothesis that tvDTH responses correlate with patient outcomes; therefore, our future directions include obtaining tvDTH responses at baseline and after initiating checkpoint inhibitor therapy in a larger and more defined cohort.
Acknowledgments
We are indebted to the patients and their families for participating in this study. We are thankful to the laboratories of Dr Douglas McNeel for providing us with pembrolizumab, Dr Paul Sondel for providing us with ipilimumab and Dr Jenny Gumperz for providing us with NSG (non-obese diabetic scid gamma) mice to perform experiments in this study.
References
Footnotes
Contributors DAL and EJ-G processed patient samples. EJ-G. performed trans vivo delayed-type hypersensitivity assays. WB and TL obtained funding for the study. TL was responsible for patient enrollment and study approval. DAL and NS performed statistical analyses. All authors contributed to data interpretation and manuscript preparation.
Funding The authors thank the University of Wisconsin Carbone Comprehensive Cancer Center for the funds to complete this project. This work is also supported in part by National Institutes of Health (NIH)/NCI P30 CA014520-UW Comprehensive Cancer Center Support and NIH Grant RO1 144AAA9541.
Competing interests TL: consultant, advisory board, Genentech, Ariad, Takeda, BMS.
Patient consent for publication Not required.
Ethics approval This study was approved by the research ethics boards of the UW-CSC (IRB 2015-1358). All patients provided written informed consent. The trial was conducted in accordance with good clinical practice guidelines.
Provenance and peer review Not commissioned; externally peer reviewed.
Data availability statement Data are available upon reasonable request. Data are available upon request to the corresponding author.