Article Text

Abstract
Background Stress-induced post-translational modifications occur during autophagy and can result in generation of new epitopes and immune recognition. One such modification is the conversion of arginine to citrulline by peptidylarginine deiminase enzymes.
Methods We used Human leukocyte antigen (HLA) transgenic mouse models to assess the immunogenicity of citrullinated peptide vaccine by cytokine Enzyme linked immunosorbant spot (ELISpot) assay. Vaccine efficacy was assessed in tumor therapy studies using HLA-matched B16 melanoma and ID8 ovarian models expressing either constitutive or interferon-gamma (IFNγ) inducible Major Histocompatibility Complex (MHC) class II (MHC-II) as represented by most human tumors. To determine the importance of CD4 T cells in tumor therapy, we analyzed the immune cell infiltrate into murine tumors using flow cytometry and performed therapy studies in the presence of CD4 and CD8 T cell depletion. We assessed the T cell repertoire to citrullinated peptides in ovarian cancer patients and healthy donors using flow cytometry.
Results The combination of citrullinated vimentin and enolase peptides (Modi-1) stimulated strong CD4 T cell responses in mice. Responses resulted in a potent anti-tumor therapy against established tumors and generated immunological memory which protected against tumor rechallenge. Depletion of CD4, but not CD8 T cells, abrogated the primary anti-tumor response as well as the memory response to tumor rechallenge. This was further reinforced by successful tumor regression being associated with an increase in tumor-infiltrating CD4 T cells and a reduction in tumor-associated myeloid suppressor cells. The anti-tumor response also relied on direct CD4 T cell recognition as only tumors expressing MHC-II were rejected. A comparison of different Toll-like receptor (TLR)-stimulating adjuvants showed that Modi-1 induced strong Th1 responses when combined with granulocyte-macrophage colony-stimulating factor (GMCSF), TLR9/TLR4, TLR9, TLR3, TLR1/2 and TLR7 agonists. Direct linkage of the TLR1/2 agonist to the peptides allowed the vaccine dose to be reduced by 10-fold to 100-fold without loss of anti-tumor activity. Furthermore, a CD4 Th1 response to the citrullinated peptides was seen in ovarian cancer patients.
Conclusions Modi-1 citrullinated peptide vaccine induces potent CD4-mediated anti-tumor responses in mouse models and a CD4 T cell repertoire is present in ovarian cancer patients to the citrullinated peptides suggesting that Modi-1 could be an effective vaccine for ovarian cancer patients.
- CD4-Positive T-Lymphocytes
- vaccination
- immunotherapy
- immunity
- cellular and adjuvants
- immunologic
This is an open access article distributed in accordance with the Creative Commons Attribution Non Commercial (CC BY-NC 4.0) license, which permits others to distribute, remix, adapt, build upon this work non-commercially, and license their derivative works on different terms, provided the original work is properly cited, appropriate credit is given, any changes made indicated, and the use is non-commercial. See http://creativecommons.org/licenses/by-nc/4.0/.
Statistics from Altmetric.com
BACKGROUND
Post-translational modifications of proteins occur under conditions of cellular stress, one such modification involves the process of citrullination—the conversion of arginine residues to citrulline (cit) is mediated by the peptidylarginine deiminase (PAD) enzymes. PAD enzymes are only activated by millimolar concentrations of calcium.1 This can occur in dead and dying cells resulting in precipitation of proteins which if it is associated with inflammation, as in rheumatoid arthritis (RA), can result in uptake by B cells and an antibody response that can result in autoimmunity.2 In contrast high levels of calcium can occur in double membrane vesicles within viable cells. Citrullination within the nucleus results in gene regulation and can have a role in tumor progression.3 4 Citrullination can also occur within autophagosomes as a result of the degradation and recycling process called autophagy which is induced in stressed cells,5 including cancer cells. In the presence of inflammation, epitopes are citrullinated and can subsequently be presented by MHC class II (MHC-II) molecules for recognition by CD4 T cells.5 CD4 T cell responses can be detected to citrullinated proteins including vimentin and enolase in autoimmune disease patients such as RA,6–9 providing further evidence that citrullinated epitopes are presented via MHC-II. We have shown that the potent immune responses unleashed in response to citrullinated vimentin can be harnessed and redirected to destroy cancer cells.10 11 Tumor recognition by the immune system depends on both citrullination and autophagy.4 10 11 This approach would benefit patients whose tumors express vimentin; this could also include mesenchymal tumors or tumors undergoing epithelial to mesenchymal transition, a process that results in an increase in tumor invasiveness, and where vimentin is one of the first proteins to be expressed.12 However, due to the heterogeneity of tumors, targeting of only one antigen has the potential to lead to the selection of HLA loss, antigen or epitope loss variants and subsequent tumor escape.13–16 HLA loss is more commonly documented than antigen loss as a result of immune pressure, although most reports are for specific MHC class I allele loss to avoid recognition by natural killer cells.13 Constitutive expression of MHC-II is largely restricted to antigen presenting cells (APCs) (dendritic cells, B cell and macrophages), thymic epithelia cells and activated T cells.17 In most other cells including epithelia, endothelia, fibroblasts and astrocytes, MHC-II can be induced by interferon-gamma (IFNγ). MHC-II expression is controlled by Class II Major Histocompatibility complex transactivator (CIITA) which is regulated by four different promoters. Promoter I is active in myeloid cells, promoter III in lymphocytes and promoter IV is essential for responsiveness to IFNγ.18 Therefore most tumors can be induced to express MHC-II unless they immune edit to prevent its expression. One of the most notable ways of inhibiting MHC-II expression is Transforming growth factor (TGF) β inhibition of CIITA but methylation of the CIITA pIV and post-transcriptional control of CIITA have also been identified.17 19 A more effective cancer vaccine could therefore be generated using multiple epitopes derived from various antigens that show restriction through multiple HLA alleles. Many tumors switch to glycolysis, even when oxygen is not limiting, in a process known as the ‘Warburg effect’.20 Glycolytic enzymes, including α-enolase, are widely expressed and are preferred targets for autophagy. We have recently shown that citrullinated α-enolase is an excellent target for anti-tumor immunity.21 T cell responses to citrullinated vimentin and enolase epitopes are documented in RA patients and we and others have shown these to be restricted through the HLA-DR*0401 (DR4) allele.6 7 10 21 In addition we have shown that responses to three peptides can also be restricted through the HLA-DP*0401 (DP4) allele22 and can stimulate strong anti-tumor immunity in HLA transgenic mice.10 21 22 Thus, combining two citrullinated vimentin peptides with a citrullinated enolase peptide should minimize tumor escape, these three peptides have been selected for inclusion in the Modi-1 vaccine. Combining epitopes can lead to dominant responses to at least one of the included epitopes with subdominant or absent responses to others.23–27 Thus, it is important to carefully select appropriate epitope combinations.
To translate a peptide vaccine into a clinical setting it is necessary to combine it with a costimulatory adjuvant. Toll-like receptor (TLR) ligand stimulation in combination with vaccination has been shown to increase vaccine efficacy, especially for cancer therapy.28 However, despite many early phase trials with new TLR ligand adjuvants only a few have been approved for use in humans. In this study we show that by combining a peptide vaccine with a TLR ligand adjuvant promotes a Th1 response, this response can be enhanced at lower peptide doses by directly linking the peptide to the TLR ligand adjuvant. The Th1 responses are capable of inducing a potent anti-tumor response in murine models of melanoma and ovarian cancer. The CD4 but not CD8 T cells were essential for the generation of the anti-tumor response, their depletion abrogated this response and a corresponding increase in CD4 tumor-infiltrating lymphocytes (TILs) was directly associated with tumor regression. In addition, human ovarian cancer patients have T cell repertoires capable of responding to these peptides thus justifying the translation of citrullinated vimentin and enolase peptide vaccine into the clinic for this cancer type.
MATERIALS AND METHODS
Cell lines and culture
The murine melanoma B16F1 cell line (ATCC-CRL-6323) and the Lewis lung carcinoma (LLC)/2 cell line (ATCC-CRL-1642) were obtained from the American Type Culture Collection (ATCC). The murine ovarian cancer line (ID8) was kindly provided by Dr K Roby at the University of Kansas Medical Center. The B16F1 cell line was cultured in RPMI medium 1640 (GIBCO/BRL) supplemented with 10% FCS, L-glutamine (2 mM) and sodium bicarbonate buffered. The ID8 and LLC/2 cell lines were cultured in DMEM (Sigma, UK) supplemented with 10% FCS. All cell lines used were certified as mycoplasma-free.
Peptides and adjuvants
Peptides vimentin aa28-49wt (RSYVTTSTRTYSLGSALRPSTS), aa28-49cit (cit-SYVTTST-cit-TYSLGSAL-cit-PSTS), aa415-433wt (LPNFSSLNLRETNLDSLPL), aa415-433cit (LPNFSSLNL-cit-ETNLDSLPL) and enolase aa241-260cit (VIGMDVAASEFF-cit-SGKYDLD) and aa241-260wt (VIGMDVAASEFFRSGKYDLD) were synthesised at >90% purity by GenScript (USA) and stored lyophilised at −80°C. Prior to use, peptides were reconstituted to the appropriate concentration in PBS (Sigma, UK). Peptides linked via the N terminus to TLR1/2 ligand AMPLIVANT were manufactured by ISA Pharmaceuticals, Netherlands.29 Adjuvants tested included murine GM-CSF (Miltenyi Biotec, Germany), TLR9 agonist CpG ODN 1826 (InvivoGen, USA), TLR4 agonist MPLA (Sigma), TLR3 agonist poly(I:C) (InvivoGen), TLR7 agonist Imiquimod (InvivoGen, USA) and TLR1/2 ligand AMPLIVANT (ISA Pharmaceuticals, Netherlands).30
Plasmids
The construction of the plasmid pVitro 2 Chimeric HLA-DR401 and the IFNγ inducible plasmid pDCGAS chimeric HLA-DR401 have been described previously.10 The HHDII plasmid pCDNA3 HHDII is described in detail elsewhere.31 Generation of the plasmid pVITRO2 human HLA-DP4 and the IFNγ-inducible plasmid pDCGAS human HLA-DP4 is described previously.22
Transfection and flow cytometry
The murine melanoma B16F1 cell line was knocked out for mouse MHC-II expression, but expresses HLA-DR4 under control of the constitutive and IFNγ inducible promoters as described previously.10 The LLC/2 lung carcinoma line expressing HLA-DP*0401 (DP4) has been described previously.22 The B16F1 cell line has been knocked out for murine MHC class I and II (B16MHKO) by Zinc Finger Technology (Sigma Aldrich, UK) and engineered to express HHDII and HLA-DP4 under a constitutive or IFNγ-inducible promoter, as described previously.22 The ID8 ovarian cancer cell line was knocked out for murine MHC class I and II (ID8MHKO) by Zinc Finger Technology (Sigma-Aldrich) was also transfected with the pVITRO2 human HLA-DP4 plasmid and selection performed using Hygromycin B (300 µg/mL). Cloning was performed by limiting dilution and expression was confirmed by flow cytometry using the anti-human HLA-DR/DP/DQ (clone WR18, Abcam, UK) PE antibodies.
Immunisation protocol
Female HLA-DR4 mice (Model # 4149, Taconic) or male and female HHDII/HLA-DP4 (DP*0401) mice (EM:02221, European Mouse Mutant Archive), aged between 8 and 12 weeks old, were used. All work was carried out at Nottingham Trent University under a Home Office approved project license. For all the studies mice were randomized into different groups and not blinded to the investigators. Peptides were dissolved in PBS and then mixed with adjuvant and delivered at a 10 nmol dose unless stated otherwise. Peptides linked to adjuvants were dissolved in PBS/DMSO and delivered as a combination at a 10 nmol dose unless stated otherwise. Adjuvants were used at the following doses; CpG ODN 1826 (TLR9) and MPLA (TLR4) at 5 µg/dose, GMCSF at 50 ng/dose, poly(I:C) (TLR3) at 10 µg/dose, imiquimod (TLR7) at 25 µg/dose and TLR1/2 agonist at 1 nmol/dose. Peptides mixed with or linked to adjuvant were injected subcutaneously at the base of the tail or intradermally unless stated otherwise. Mice were immunized on days 0, 7 and 14, spleens removed and splenocytes harvested for analysis at day 20.
For anti-tumor therapy experiments mice were challenged with 2.5×104 B16 DR4 cells, 4×105 B16 HHDII DP4 cells, 4×105 ID8 DP4 cells (in Matrigel (Corning Life Sciences, UK)), 5×104 B16 inducible DR4 cells or 1×105 B16 inducible DP4 cells subcutaneously on the right flank 3 days prior to primary immunization and were then immunized as above unless stated otherwise. In studies using anti CD4 (anti-human CD4 clone OKT4, BioXcell) and anti CD8 (anti-mouse CD8 clone 2.43, BioXcell) depletion antibodies, mice were dosed with 500 µg antibody (intraperitoneally) plus peptide on day 4 followed by 300 µg antibody plus peptide on days 8 and 11. For the tumor rechallenge studies, tumor-free mice were implanted with 1×105 B16 inducible DP4 cells or 2.5×106 LLC/2 DP4 cells on days 50 or 58 without any further immunization. Tumor growth was monitored at 3 to 4 day intervals and mice were humanely euthanized once the tumor reached 10 to 15 mm in diameter. For tumor rechallenge studies using depletion antibodies, 500 µg antibodies were administered 1 day prior to the rechallenge and 300 µg at 2 and 6 days post tumor rechallenge.
For the TIL analysis and regression studies, mice were challenged with 1×105 B16 inducible HLA-DP4 cells subcutaneously on the right flank, once tumors reached 8 to 12 mm in diameter mice were immunized with a single dose of 10 nmol of each peptide in CpG/MPLA adjuvants. Tumor size was monitored and were excised after 4 days for analysis. Tumor size was calculated as follows: volume = (π/6) x (width x length)2. Regression/progression was calculated as tumor volume change from day of vaccination to analysis point at day 4 post vaccination.
Ex vivo ELISpot assay
ELISpot assays were performed using murine IFNγ capture and detection reagents according to the manufacturer’s instructions (Mabtech). In brief, the IFNγ-specific antibody were coated onto wells of 96-well Immobilin-P plates. Synthetic peptides (GenScript) were used at 10 µg/mL and diluted in culture media (RPMI medium 1640 (GIBCO/BRL) supplemented with 10% FCS (Sigma), 2 mM L-glutamine (Sigma) and sodium bicarbonate buffered with additional 20 mM HEPES (Sigma) and 50 µM 2-mercaptoethanol (2ME, Thermo Fisher) and 5×105/well splenocytes were added in quadruplicate wells, plates were incubated for 40 hours at 37 °C/5% CO2. Cells with culture media only were added as negative control and cells cultured with 5 µg/mL lipopolysaccharide (Sigma) were used as a positive control. Tumor cells were used either untreated, or stress induced by treatment with cold 0.1M citrate buffer pH3 for 2 minutes followed by 16 hours incubation in serum free media and 5×104 cells were added to each well. After incubation, captured IFNγ was detected by biotinylated-specific IFNγ antibody and developed using streptavidin alkaline phosphatase and chromogenic substrate. Spots were analyzed and counted using an automated plate reader (Cellular Technologies Ltd). Results were represented as spots/million splenocytes. Our studies in this report make use of a 40 hours incubation time for the cytokine measurement which is longer than most reported assays. We can show that there is little difference in number of responding cells and total cytokine release between the time points analyzed here and earlier time points traditionally examined (online supplementary figure 1A).
Supplemental material
TIL analysis
Tumors were removed from mice and mechanically disaggregated in 2 mL of culture medium (RPMI 10% FCS, 20 mM HEPES, 2 mM L-glutamine, Penicillin (100 U/mL, Sigma, UK) Streptomycin (100 µg/mL, Sigma, UK), 2ME (10-5M). Cells were transferred to 15 mL tubes and large debris allowed to settle for 5 min. Supernatant was transferred to a new tube, cells were counted and resuspended at approximately 1×106/mL in culture media, 0.5×106 to 1×106 cells were removed, washed with PBS and stained with cell surface markers (CD8 VioGreen clone REA601 (Miltenyi Biotec), CD45 efluor450 clone 30-F11 (Thermo Fisher), human CD4 APCefluor780 clone RPA-T4 (Thermo Fisher), CD11b VioGreen clone REA592 (Miltenyi Biotec), Gr-1 FITC clone 1A8 (Thermo Fisher)). Cells were then resuspended in fixation buffer (Thermo Fisher) prior to running on the MACSQuant (Miltenyi Biotec) flow cytometer.
Patient samples
The anonymized details of donors and patients are given in online supplementary table 1.
Supplemental material
Proliferation assays on human PBMCs
Peripheral blood (approximately 50 mL) was drawn into lithium heparin tubes (Becton Dickinson). Samples were maintained at room temperature and processed immediately following venepuncture. Peripheral blood mononuclear cells (PBMCs) were isolated by density gradient centrifugation using Histopaque-1077 (Sigma, UK). Proliferation assays were performed immediately after PBMC isolation. The median number of PBMCs routinely isolated from healthy donors was 1.04×106 PBMC/mL whole blood (range: 0.6×106 to 1.48×106/ mL). The median viability as assessed by trypan blue exclusion was 93% (range 90% to 95%). The median number of PBMCs routinely derived from cancer patients was 1.2×106 PBMC/mL whole blood (range: 0.44×106 to 2.5×106/ mL). The median viability as assessed by trypan blue exclusion was 90% (range 76% to 97%). For CD25 depletions, PBMCs were processed as above and then immediately depleted of CD25+ cells using anti-CD25 microbeads and MACS Cell Separation Columns (Miltenyi).
Freshly isolated CD25 depleted PBMCs were loaded with carboxyfluorescein succinimidyl ester (CFSE) (Thermo Fisher). Briefly, a 50 µM CFSE stock solution was prepared in warm PBS from a master solution of 5 mM in DMSO. For loading, CFSE was rapidly added to PBMCs (5×106 cells/mL loading buffer (PBS with 5% v/v heat inactivated FCS)) to achieve a final concentration of 5 µM. PBMCs were incubated at room temperature in the dark for 5 min, after which non-incorporated CFSE was removed by washing the cells twice with excess (×10 v/v volumes) of loading buffer (300 g x 10 min). Cells were made up in complete media to 1.5×106/mL and plated in a 24 well plate (2 mL/well), cells were stimulated with PHA (positive control, final concentration 10 µg/mL) or peptides (10 µg/mL), or were unstimulated (negative control, containing DMSO vehicle).
After 7 to 11 days post culture set up, 500 µL of cells were removed, treated with Brefeldin A for 4 hours, washed in PBS and stained with 1:50 dilution of anti-CD4 (PE-Cy5, clone RPA-T4, Thermo Fisher), anti-CD8 efluor 450, clone RPA-T8, Thermo Fisher) and anti-CD134 (PE-Cy7, Clone REA621, Miltenyi). Cells were washed, fixed and permeabilised using intracellular fixation/permeablization buffers (Thermo Fisher) according to the manufacturer’s instructions. Intracellular staining for cytokines was performed using a 1:50 dilution of anti-IFNγ (clone 4S.B3, Thermo Fisher) or anti-Granzyme B (PE, Clone GB11, Thermo Fisher). Stained samples were acquired on a MACSQuant 10 flow cytometer equipped with MACSQuant software V.2.8.168.16380, the stained unstimulated controls were used to determine suitable gates.
Statistical analysis
Comparative analysis of the human proliferation assay results was performed by applying paired Student's two-tailed t-test and the human cytokine analysis performed using unpaired multiple t-tests with values of p calculated accordingly. Comparative analysis of the ELISpot results were performed by applying ordinary one-way analysis of variance with Sidak’s multiple comparisons test and values of p calculated accordingly. Comparison of tumor survival was assessed by log-rank test using GraphPad Prism software. The association between tumor growth and TIL infiltration was assessed using linear regression. For each study, the percentage of cells in each tumor staining positive for the CD45, CD4 and CD8 markers was determined from the average of at least three replicate staining tubes per sample. P values <0.05 were considered statistically significant.
RESULTS
Citrullinated vimentin and enolase peptides can be combined into a single vaccine and mediate efficient tumor therapy
Clinically, targeting of only one antigen has the potential to lead to the selection of HLA loss, antigen or epitope loss variants and subsequent tumor escape. A more effective cancer vaccine could be generated targeting multiple HLA alleles and using multiple epitopes derived from various antigens however the latter can lead to immunodominance of one of the epitopes. Thus we compared immunization of HLA-transgenic mice with single or combinations of citrullinated peptides. Immunization of mice with individual peptides led to high-frequency epitope-specific IFNγ responses to citrullinated aa28-49 vimentin, aa415-433 vimentin and aa241-260 enolase peptides. On combination of the three peptides into the same vaccine (Modi-1) similar high-frequency peptide-specific responses were observed in both HLA-DR4 (figure 1Ai) and HLA-DP4 (figure 1Aii) mice, suggesting there was no immune competition between these three epitopes. Indeed, the responses to some epitopes were significantly improved within the combination (Eno241cit p=0.0001 in HLA-DR4 mice and vim415cit p=0.0001 in HLA-DP4 mice). The combination of three Modi-1 peptides shows no immune competition therefore these are a candidate for a vaccine to target both multiple HLA alleles and multiple antigens and thus broadens the clinical applicability of this vaccine. In addition to showing little immune competition between the three epitopes we demonstrate that responses induced are specific to the citrullinated peptides with no cross reactivity to the native (wild type (wt)) sequences (figure 1Ai,ii). Immune responses to the three epitopes in both HLA-DR4 and DP4 mice have previously been shown to be CD4 mediated10 21 22 and this is also confirmed in this study (online supplementary figure 1B). In addition to the specific detection of IFNγ responses to the citrullinated peptides we also demonstrate here the specific release of granzyme B (figure 1Aiii). Direct tumor recognition is also confirmed in vitro with splenocytes from immunized mice showing both peptide-specific IFNγ responses and responses to stressed tumor cells that was not seen with splenocytes from naïve mice (figure 1Aiv).
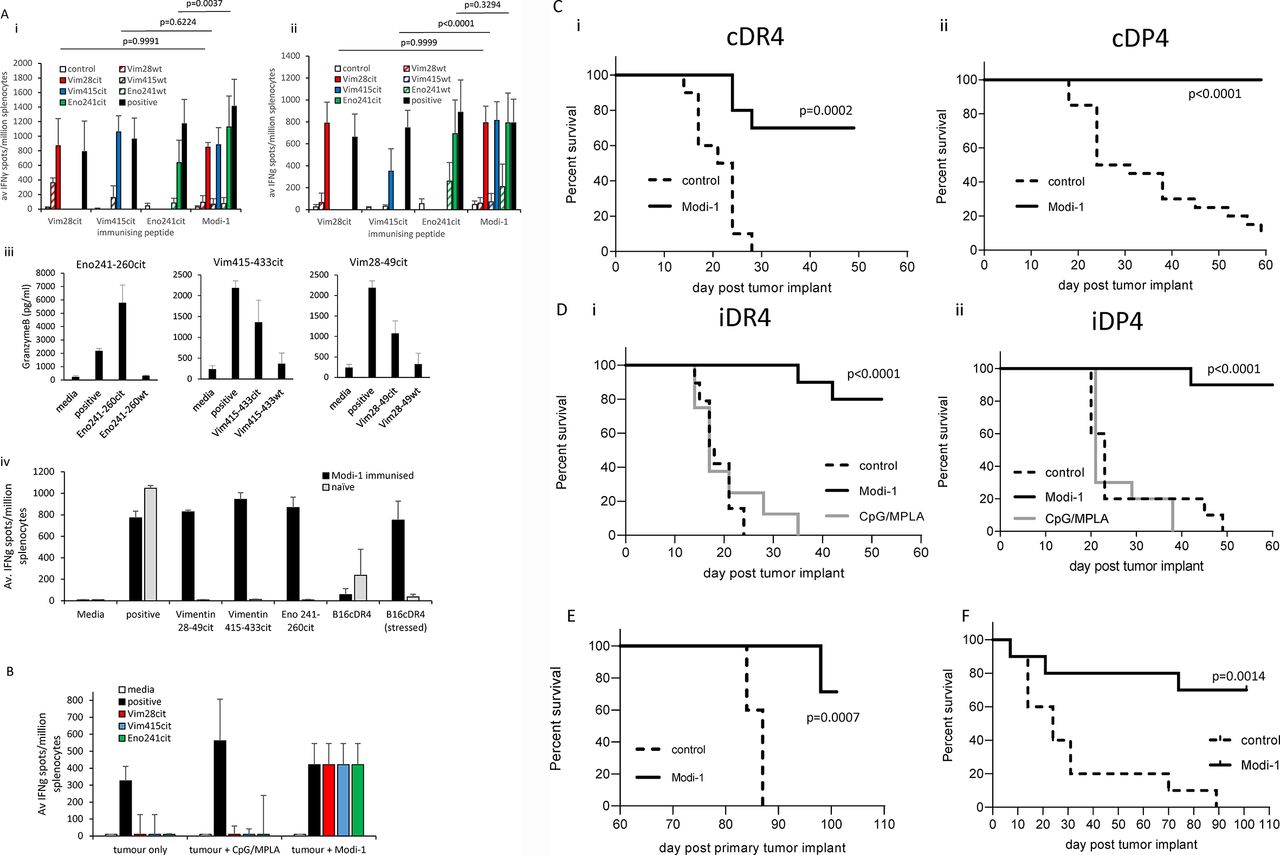
Combination of citrullinated vimentin and enolase peptides stimulates a CD4 T cell response that mediates anti-tumor therapy. HLA-DR4 (Ai) or HLA-DP4(Aii) transgenic mice were immunized with single or a combination (Modi-1) of citrullinated Vim28-49, Vim415-433 and Eno241-260 peptides (10 nmol each) in CpG/MPLA, immune responses specific to the citrullinated or wild type peptides were monitored by IFNγ ELISpot assay and compared with control. HLA-DP4 mice immunized with the combination vaccine were analyzed for granzyme B responses specific to the citrullinated peptides by ELISA (Aiii) and splenocytes from HLA-DR4-immunized mice were analyzed for IFNγ responses to B16cDR4 tumor cells in vitro (Aiv). Results are representative of at least two independent experiments in which n=3 mice per group. B, HLA-DP4 mice were implanted with B16 cells expressing DP4 under the control of an IFNγ inducible promoter followed by vaccination with Modi-1 in CpG/MPLA or CpG/MPLA adjuvant and immune responses specific to citrullinated peptides analyzed by IFNγ ELISpot assay. Responses normalized against background control. HLA-DR4 (i) or DP4 (ii) transgenic mice were implanted with B16 cells either constitutively expressing DP4 or DR4 (Ci and Cii) or under the control of an IFNγ inducible promoter (Di and Dii). Three days later mice were immunized with a single immunization (Dii) or multiple immunizations (Ci, Cii and Di) of Modi-1 peptides (10 nmol each) or CpG/MPLA adjuvant only, tumor growth and survival was monitored. On day 58 surviving tumor free HLA-DP4 transgenic mice from the Modi-1 tumor therapy study were rechallenged with a second dose of B16 cells expressing DP4 under the control of an IFNγ inducible promoter (E), tumor-free survival was monitored compared with new unimmunized control mice. HLA-DP4 transgenic mice were implanted with ID8 cells constitutively expressing HLA-DP4 (F), 4, 11 and 18 days later mice were immunized with the Modi-1 peptides, tumor growth and survival were monitored. n=10 mice per group. IFNγ,interferon-gamma.
To investigate if tumor bearing mice either with or without immunization showed Modi-1 specific responses, mice bearing tumors were immunized with Modi-1. Untreated tumor bearing mice or those receiving only CpG/MPLA adjuvant show no specific responses to Modi-1 peptides (figure 1B). In contrast tumor-bearing mice expressed strong responses to all three Modi-1 epitopes. The three Modi-1 peptides in combination were assessed for their ability to mediate tumor therapy. HLA-DR4 or HLA-DP4 mice were challenged with B16MHKO-DR4/DP4 tumor, as appropriate, and subsequently vaccinated with the Modi-1 peptides plus adjuvant. Transgenic mice showed a good anti-tumor response when peptides were combined with TLR9/TLR4 ligands (figure 1C). The difference in tumor growth reflects the clonal variation in these lines. B16 tumors are aneuploid and show great clonal variation, in selecting for the mouse MHC knock out and human HLA transgenic lines clonal variation resulting in differing growth rates can occur. As many tumors do not constitutively express high levels of MHC-II molecules, although most are responsive to MHC-II induction by IFNγ, a model was engineered that expressed HLA-DR4 (iDR4) or HLA-DP4 (iDP4) under the control of an IFNγ inducible promoter (online supplementary figure 2A–C). Analysis of Modi-1 responses induced in the presence of TLR9/TLR4 ligands showed efficient tumor therapy even when expression of MHC class II on the tumor is reliant on induction by IFNγ in vivo (figure 1D). To determine if the tumor effect was specific to the citrullinated peptides, mice were challenged with tumor and immunized with the murine unmodified (wt) or citrullinated peptide sequences in combination with TLR9/TLR4 adjuvant or the adjuvant alone. Citrullinated peptides showed good anti-tumor responses but immunization with adjuvant alone or with the native peptide sequences provided no tumor therapy (online supplementary figure 3). To assess the longevity of the immunity generated by the Modi-1 vaccine, surviving tumor-free mice were rechallenged with the same tumor line and tumor growth monitored. Those mice that were vaccinated with Modi-1 and were tumor-free following the first tumor challenge were rechallenged with the same tumor. Of these 71% survived; this was significantly higher than the control group (p=0.0007). This increased survival was generated without any further vaccination suggesting the establishment of an immunological memory response (figure 1E).
In addition to the B16MHKO-DR4/DP4 melanoma models, we next looked at a murine ovarian tumor line (ID8MHKO-DP4) in the HLA-DP4 (cDP4) mouse model, this line expresses both vimentin and enolase (online supplementary figure 4). Those mice vaccinated with Modi-1 showed significantly higher survival than the control group (p=0.0014), indicating the potential of Modi-1 to improve survival in this indication (figure 1F).
Tumor regression mediated by Modi-1 peptide vaccination correlates with increase in CD4 T cell infiltrate and a decrease in myeloid suppressor cell population
HLA-DP4 mice with a large tumor burden were subsequently vaccinated with Modi-1 or adjuvant only and tumor infiltrates were analyzed. To account for variation in tumor growth between mice, they were vaccinated when tumor diameters reached 8 to 12 mm and tumor infiltrates analyzed after 4 days. Despite Modi-1 vaccinated mice showing slightly larger tumors at vaccination point compared with the control group (p=0.0030), comparison of tumor volume pre-immunization and at day 4 post-immunization show regression in the Modi-1-immunized mice (figure 2Ai). Tumors from Modi-1-immunized mice showed evidence of rapid tumor regression within 4 days of a single immunization which was not seen in control mice receiving adjuvant only (figure 2Aii). Seven out of 10 Modi-1-immunized mice showed evidence of tumor shrinkage, measured as tumor volume change, with the remaining 3/10 showing stable disease whereas mice treated with the adjuvant alone showed 7/9 mice with tumor progression and 2/9 with stable disease (figure 2Aiii). Tumors were removed at this time point and the infiltrating immune cells were analyzed based on the expression of CD45; the proportion of CD45+ cells that were either CD4 or CD8 T cells or tumor associated MDSCs (CD11b+Gr1+) was determined. In Modi-1-immunized mice with regressing tumors there was no significant difference in the percentage of CD45+ cells in the tumors when compared with the control group (figure 2B). However, the regression of tumors in the Modi-1-immunized mice correlated with an increase in the percentage of tumor-infiltrating CD4 T cells, suggesting that CD4 T cells may play a key role in tumor regression (figure 2C). Tumors from mice immunized with Modi-1 showed a decrease in tumor-infiltrating MDSCs (myeloid-derived suppressor cells),32 that correlated with tumor regression (figure 2D).
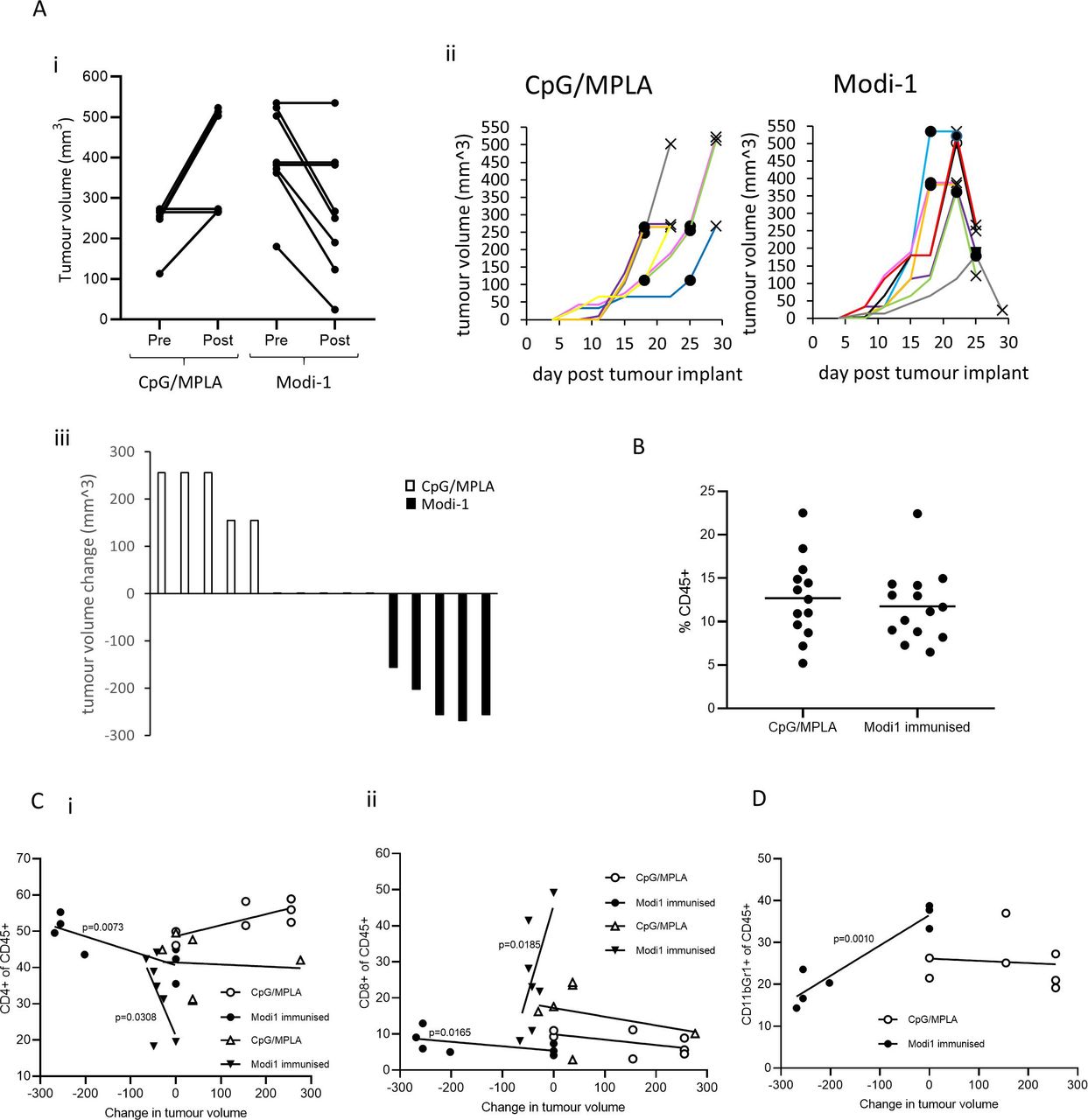
Modi-1 vaccination causes rapid tumor regression which is associated with increased CD4 T cell infiltration and a decrease in MDSCs. Mice were immunized when tumors reached 8 to 12 mm diameter with either a single dose of Modi-1 peptides in combination with CpG/MPLA or with CpG/MPLA only and tumor volume was monitored (A). Tumor volume at vaccination day compared with day 4 post-vaccination (i) and tumor growth over time (ii) circles (·) denote the day of immunization for each mouse and crosses (X) indicate the day of tumor analysis. Waterfall plot of tumor volume change between immunization and termination (iii). Data from a representative study is shown. Dissociated tumors were analyzed by flow cytometry to determine the percentage of CD45+ cells (B), percentage of CD4 (i) or CD8 (ii) T cells among CD45+ cells (C) and percentage of MDSCs (CD11b+, Gr1+) among CD45+ cells (D). The percentage of CD4, CD8 or MDSCs among the CD45 population were correlated to tumor volume change (C,D). Data shown in D is from two independent studies (circles and triangles) in which n=at least 6. MDSCs,myeloid-derived suppressor cells.
CD4 T cells drive Modi-1 vaccine mediated tumor therapy
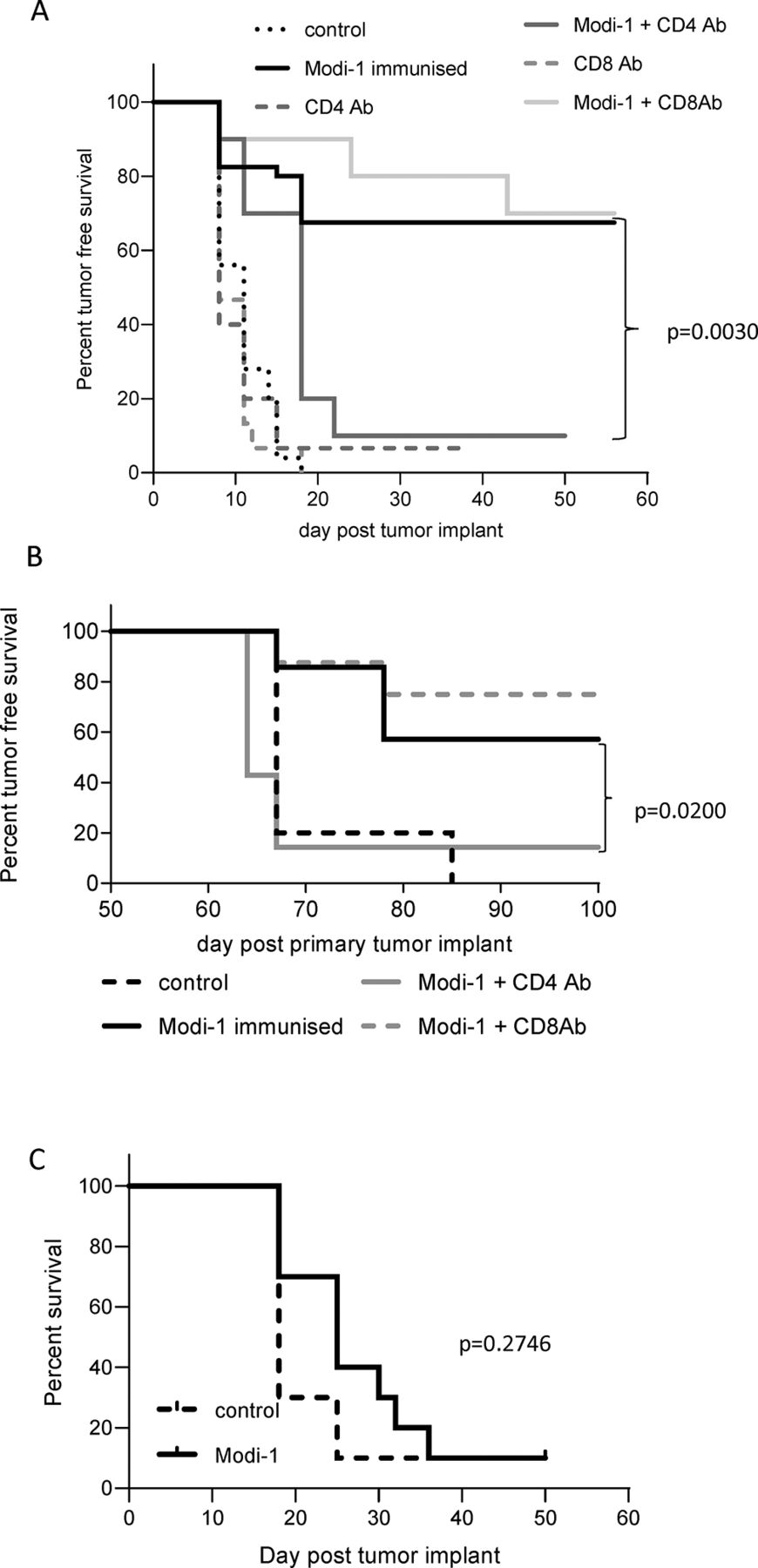
In addition to the infiltrate analysis to determine the role of CD4 and CD8 T cells in tumor therapy we performed in vivo depletion of CD4 or CD8 T cells in mice at the same time as vaccinating with Modi-1. Both antibodies were shown to deplete CD4 or CD8 T cells in both the blood and tumor samples from the B16MHKO-DP4 melanoma mouse model (online supplementary figure 5). Depletion of CD4 T cells abrogated the anti-tumor effect mediated by the vaccine (p=0.0030), whereas the depletion of CD8 T cells had no significant effect (p=0.80) (figure 3A). Since the Modi-1 vaccine generates immunological memory that can protect against subsequent tumor rechallenge we next examined the role of CD4 and CD8 T cells in the memory response by measuring tumor-free survival following tumor rechallenge. The depletion of CD4 T cells at the time of tumor rechallenge resulted in a loss of the Modi-1 protection, as shown by a significant reduction in tumor-free survival (p=0.02), whereas the depletion of CD8 T cells did not show any significant effect (p=0.512) (figure 3B). Therefore, we can conclude that the CD4 T cell responses play a vital role in both the initiation of anti-tumor therapy and the protection against rechallenge.

Anti-tumor therapy is dependent on the presence of CD4 T cells. HLA-DP4 transgenic mice were implanted with B16 cells expressing DP4 under the control of an IFNγ-inducible promoter and immunized with Modi-1 peptides in CpG/MPLA on days 4, 8 and 11 in the presence of CD4 or CD8 depleting antibodies (A). Mice who remained tumor-free following Modi-1 immunization were rechallenged on day 58 with B16 cells expressing DP4 under the control of an IFNγ-inducible promoter, in the presence of either CD4 or CD8 depleting antibodies (B). HLA-DP4 transgenic mice were challenged with B16 cells expressing appropriate MHC class I (HHDII) but lacking MHC class II expression, 4, 11 and 18 days later mice were immunized with Modi-1 (C). Tumor growth and survival were monitored. n>8 mice per group. IFNγ,interferon-gamma.
CD4 T cells can play a role in tumor therapy via both indirect and direct mechanisms. Indirect mechanisms such as recruitment of other immune cells or cytokine production do not require expression of MHC-II by the tumor, whereas direct recognition does. Therefore we next investigated the effect of the vaccine in a model where the tumor was unable to express MHC-II. Despite infiltrating APCs still being able to present antigen via the relevant MHC-II in this model, the anti-tumor effect was lost (p=0.2746), indicating a role in the direct recognition of the tumor by CD4 T cells (figure 3C).
Th1 responses to Modi-1 peptides can be induced in combination with a variety of TLR ligand adjuvants
Previous vaccination studies have indicated that a more efficient proinflammatory response can be induced by the inclusion of an appropriate adjuvant providing costimulation. To determine the best adjuvant to combine with the Modi-1 peptides, the immune responses in HLA-DR4 and HLA-DP4 mice were analyzed after immunization with citrullinated vimentin and enolase peptides in combination with an adjuvant: CpG (TLR9), MPLA (TLR4), CpG/MPLA (TLR9/TLR4), GMCSF, imiquimod (TLR7), poly I:C (TLR3) or TLR1/2 ligand (AMPLIVANT). In HLA-DR4 mice a high frequency of specific T cells were detected in response to all citrullinated peptides when combined with either TLR9/TLR4, TLR3, GM-CSF and TLR2 agonists, whereas combination with TLR4 agonist alone induced only minimal IFNγ responses (figure 4Ai). The lack of response with the TLR4 agonist suggests the responses seen with the TLR9/TLR4 combination are a result of the TLR9 agonist. Some of these adjuvant combinations were also tested in the HLA-DP4 mouse model which showed a similar pattern of responses including a response with the TLR9 agonist in addition to the TLR9/TLR4 combination (figure 4Aii). The use of the TLR1/2 adjuvant showed higher frequency responses compared with GMCSF (p<0.0001), TLR3 (p=0.0036) and TLR7 (p=0.0055) for aa415-433cit vimentin peptide, GMCSF (p=0.0002) for aa28-49cit vimentin peptide and GMCSF (p=0.0074) and TLR7 (p=0.0038) for aa241-260cit enolase in HLA-DR4 mice. In HLA-DP4 model TLR1/2 adjuvant showed higher frequency responses compared with TLR3 and TLR7 adjuvants (p<0.02) for all peptides; this adjuvant was therefore selected for further investigation.

Citrullinated vimentin and enolase peptides stimulate CD4 T cell responses when given in combination with TLR ligands, linking the peptides to adjuvant promotes a stronger immune response also resulting in an anti-tumor response. HLA-DR4 (i) or DP4 (ii) transgenic mice were immunized with 1 to 10 nmol dose of Modi-1 peptides given either individually or in combination with various adjuvants, immune responses specific to the Modi-1 peptides were monitored in an IFNγ ELISpot assay (A). Results are shown from at least two independent experiments in which n=3 mice per group. HLA-DR4 (i) or HLA-DP4 (ii) transgenic mice were immunized with decreasing doses of Modi-1 peptides mixed with TLR1/2 or linked to TLR1/2 adjuvant (B). Immune responses specific to the Modi-1 peptides were monitored in an IFNγ ELISpot assay. Results are representative of at least two independent experiments in which n=3 mice per group. HLA-DR4 (i) or HLA-DP4 (ii) transgenic mice were implanted with B16 cells expressing DP4 or DR4 under the control of an IFNγ-inducible promoter, 3 days later mice were immunized with 0.1 nmol Modi-1 peptides linked to TLR1/2 adjuvant (C). Tumor growth and survival was monitored, n=10 mice per group. HLA-DP4 mice who remained tumor-free following Modi-1 immunization were rechallenged on day 50 with B16 cells expressing HLA-DP4 under the control of an IFNγ-inducible promoter, survival was monitored (D) or LLC/2 DP4 cells and tumor growth assessed at day 60 (E). IFNγ,interferon-gamma; ND, notdetermined; TLR, Toll-like receptor.
Linkage of peptide to TLR agonist enhances immune responses and promotes tumor therapy with lower dose vaccine
Previous reports suggest that the direct conjugation of peptides to TLR ligands enhances immune responses by delivering the adjuvant and the antigen to the same APC.33 34 We therefore investigated whether a lower dose of Modi-1 peptides linked with the TLR1/2 ligand AMPLIVANT (online supplementary figure 6) could generate a comparable immune response to that seen with the higher peptide doses previously tested (1 to 10 nmol). HLA-DR4 (figure 4Bi) and HLA-DP4 (figure 4Bii) mice were immunized with decreasing doses of the Modi-1 peptides given in combination or linked to the TLR1/2 ligand AMPLIVANT. In both mouse models good immune responses were still observed at concentrations as low as 0.01 nmol when linked to TLR1/2 ligand AMPLIVANT, however, at this concentration these responses were lost when peptides were mixed with TLR1/2 ligand AMPLIVANT. These results demonstrate that by linking the vimentin and enolase peptides to TLR1/2 ligand AMPLIVANT (Modi-1) a comparable specific immune response can be generated in mice at lower peptide doses, indicating that it has the potential to be dose sparing in the clinic.
We next investigated whether a lower dose of Modi-1 (0.1 nmol) could mediate an anti-tumor effect in an aggressive tumor model in mice. HLA-DR4 (figure 4Ci) or DP4 (figure 4Cii) transgenic mice were implanted with the melanoma B16MHKO cell line that expressed inducible DR4 or DP4 and immunized with 0.1 nmol Modi-1. Mice immunized with Modi-1 showed significant survival in both DR4 model when compared with unimmunized mice (p=0.0049) or those that received adjuvant alone (p=0.0221) and in DP4 model compared with unimmunized mice (p=0.0002). Mice surviving the primary melanoma B16 tumor also rejected a subsequent rechallenge with the same tumor (p<0.0001) (figure 4D). Survivors of primary melanoma challenge were also assessed for protection against rechallenge with the LLC/2 lung tumor cell line, known to express vimentin and enolase and constitutively expressing HLA-DP4. Without any further vaccination, the previously Modi-1 immunized and challenged mice showed a significant delay in tumor growth (p=0.0007) when rechallenged with a different tumor when compared with unimmunized mice (figure 4E).
Ovarian cancer patients show a repertoire of CD4 T cells that respond to citrullinated peptides
Self-antigen-specific CD4 responses are known to be regulated by Tregs (regulatory T cells) in vivo and in vitro,35 36 therefore to determine the available repertoire specific to Modi-1 peptides we sought to examine responses in the absence of Tregs. We have already shown that a repertoire of T cells exist in healthy donors that can respond to these vimentin and enolase-derived citrullinated peptides.21 22 We next investigated whether cancer patients also have a repertoire of T cells that respond to these peptides. We tested the T cell response to peptide stimulation in PBMCs from 17 ovarian cancer patients (online supplementary table 1B). Ten of the patients (58%) showed a CD4 T cell proliferative response that was above twice the background and greater than 1% for at least one of the Modi-1 peptides (figure 5Ai,ii). However, only two cancer patients (OV18 and OV19) responded to both citrullinated vimentin and enolase. The proliferative responses in cancer patients were predominantly CD4 T cell mediated as shown in example plots from donors OV18 and OV19 (figure 5B). Comparison of responses between cancer patients and healthy donors reveals no significant difference in responses to any of the three peptides tested (figure 5C). Analysis of the HLA restrictions of patients and healthy donors reveals most responders to express either HLA-DR4 or HLA-DP4 allele suggesting these are likely restriction elements in the human donors (online supplementary table 1). However, HLA typing was not available for all patients and not all healthy responders expressed one of these two alleles. For example healthy donor BD0011 was neither DR4 nor DP4 positive implying the presence of another restricting allele. Using the predictive algorithm IEDB for binding affinity of the Modi-1 sequences to MHC-II alleles present in BD0011 shows strong predictions for HLA-DQ6, HLA-DR13 and HLA-DP18 where the arginine/citrulline lies within the predicted core binding region. Although this does not provide definitive evidence for binding of the citrullinated peptides it suggests that other MHC-II alleles may be able to present these peptides (online supplementary table 2).

Responses to Modi-1 peptides in cancer patients and healthy donors. PBMCs from cancer patients stimulated ex vivo with Modi-1 peptides, the proliferative response was measured (A). Results are shown as a percentage of proliferating CD4 T cells shown for each peptide (i) and each patient (ii). Example flow cytometry dot plots showing proliferation of CD4 T cells from cancer patients in response to Modi-1 peptides (B) with values displayed representing the percentage of proliferating CD4 T cells. Comparison of proliferating CD4 T cells in healthy donors and cancer patients (C) in response to each of the Modi-1 peptides. Data normalized against background control. PBMCs, peripheral blood mononuclear cells.
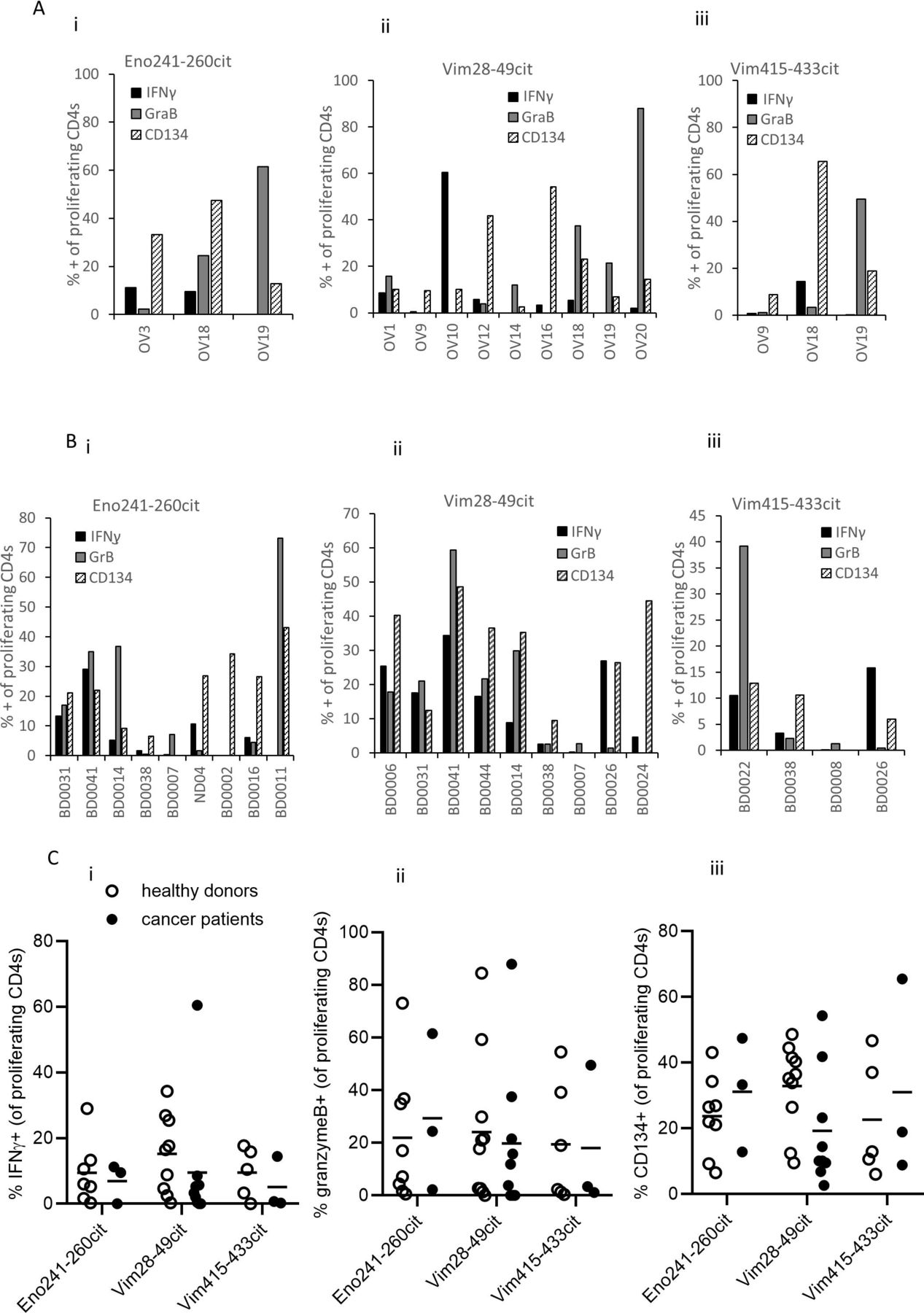
To further characterize the responding T cells the expression of IFNγ, granzyme B and CD134 was measured on the proliferating CD4 T cells. Examples of staining is shown in online supplementary figures 7 and 8. Of the three cancer patients proliferating to enolase aa241-260cit, cells show up to 45% expression of CD134, lower expression of IFNγ and variable expression of granzyme B (figure 6Ai). All cultures proliferating to vimentin aa28-49cit showed expression of CD134, IFNγ and granzyme B, although these were variable between patients (figure 6Aii). Similarly all cultures proliferating to vimentin aa415-433cit expressed CD134, however only one of three showed IFNγ staining and two of three granzyme B staining (figure 6Aiii). Despite varying levels of staining, these results suggest that the responding T cells have a cytotoxic Th1 phenotype. Healthy donor responses show a similar phenotype to responses from the cancer patients. All cultures except one from healthy donors proliferating to enolase aa241-260cit showed expression CD134 to a similar level to that seen in the cancer patients (figure 6Bi). Variable IFNγ and granzyme B staining was also seen between donors. All cultures except one from healthy donors proliferating to vimentin aa28-49cit expressed CD134 but similar to enolase aa241-260cit responses the staining of IFNγ and granzyme B was variable (figure 6Bii). Three of four cultures from healthy donors proliferating to vimentin aa415-433cit expressed CD134 and three of four also showed staining for granzyme B although this was variable. Variable IFNγ staining was also seen in three of four cultures. Comparison of all peptide-specific responses in healthy donors and the cancer patients showed no significant difference in the expression of IFNγ, granzyme B or CD134 suggesting that the T cell repertoire in cancer patients in this study have a similar functional T cell repertoire to healthy individuals for the markers analyzed (figure 6C).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Characterizing the immune response to Modi-1 peptides in PBMCs from cancer patients and healthy donors. PBMCs from cancer patients (A) and healthy donors (B) were stimulated with the individual Modi-1 peptides, the expression of IFNγ, granzyme B and CD134 was determined on proliferating CD4 T cells in response to citrullinated enolase 241–260 (i), vimentin 28–49 (ii), or vimentin 415–433 (iii). Results shown as percentage proliferating CD4 T cells. Comparison of IFNγ (i), granzyme B (i) and CD134 (iii) on proliferating CD4 T cells from healthy donors and cancer patients (C) in response to each of the citrullinated peptides. IFNγ,interferon-gamma; PBMCs, peripheral blood mononuclearcells.
DISCUSSION AND CONCLUSIONS
Many cancer vaccines have been focused on the stimulation of CD8 T cell responses and their role in tumor therapy rather than CD4 responses. In this study we have shown that mouse tumors present citrullinated vimentin and enolase epitopes that are targets for cytotoxic CD4 T cell responses.10 21 More recent findings suggest the use of longer synthetic peptides as vaccines yields better immune responses than minimal epitope peptides with induction of both CD8 and CD4 responses.37–39 CD4 T cells have been shown to play an important role in cancer therapy and can either directly mediate the destruction of established tumors40 41 and/or indirectly by their interaction with APCs which induces the release of interferons that then upregulate MHC-I/II and stimulate the release of proinflammatory chemokines to promote further immune infiltration.42 43
We have shown that a therapeutic peptide vaccine that targets modified self-antigens can induce a Th1 immune response that is specific to the modified antigen with no reactivity to the native sequences and mediated a potent anti-tumor response in a melanoma mouse model. This response seems to be mediated by CD4 T cells as blockade of CD4 T cells abrogated the response in vitro and the depletion of CD4 T cells in vivo diminished the anti-tumor response whereas the depletion of CD8 T cell had no impact. A role for direct CD4 T cell-mediated recognition of tumor cells was demonstrated in vitro by splenocytes from Modi-1-immunized mice but not naive mice. The loss of the in vivo anti-tumor response in the absence of MHC-II expression by the tumor also supports the role of direct CD4 T cell recognition of the tumor cells. Reduction in MDSC in regressing tumors suggest that the CD4s may be also mediating indirect effects allowing repolarization of the cells but this alone did not seem to have a significant anti-tumor effects as there was not anti-tumor immunity against tumors lacking MHC-II expression.
The CD4 repertoire to self-antigens is known to be highly tolerized and as a result there are few reports of vaccine-induced CD4 T cell responses to self-antigens.44 However, an exception to this is the discovery of a number of CD4 T cells recognizing tumor-specific mutations or neo-epitopes. These neo-epitopes are patient specific and can be successfully identified and included in a personalized cancer vaccine or immunotherapy strategies.45–47 In addition to neo-antigen-specific CD4 T cells, CD4 T cell responses to post-translational modifications such as citrullination provide another group of candidates to target for inducing anti-tumor immunity. Citrullination is associated with autoimmunity, in particularly RA, mostly through the generation of anti-citrulline protein antibodies (ACPAs). RA is known to be induced not by T cells alone but by the combination with joint erosion, antibody responses and inflammation. Evidence to support this was obtained from studies in HLA-DR4 transgenic mice, a strain known to be susceptible to RA, where no disease was observed with T cells alone.48 In addition, this study and those from others have not shown any autoimmune effects in susceptible mouse models after vaccination with citrullinated peptides suggesting other factors in addition to citrulline-specific CD4 T cells are required for onset of autoimmunity.9 This does not exclude the possibility that autoimmunity may be exacerbated in patients that already have the disease. Therefore signs of autoimmunity should be monitored and patients with known underlying autoimmunity excluded from any future clinical trial.
In this study we measured the CD4 T cell response to three citrullinated peptides derived from vimentin and enolase. We chose to combine the three epitopes based on our previous findings that they provided efficient tumor therapy individually.10 21 As the anti-tumor responses to individual peptides was so strong we did not expect to see a synergistic effect in the mouse model but may see this in cancer patients who are much harder to treat than animal models. It was however important to show that the peptides did not compete, thereby reducing the anti-tumor response. Such competition is often seen in viral infections where immunodominant CD4 T cell responses predominate;49 our results show that there is no apparent immunodominance between the citrullinated epitopes. We have previously shown that although the citrullinated epitopes bind more strongly to MHC-II than the wild type epitopes they still only have a moderate binding affinity.22 In the context of autoimmunity, T cells targeting self-antigens have been shown to have a low binding affinity to peptide-MHC-II complexes, despite the editing effect of HLA-DM, these peptides are often more resistant to proteolytic cleavage allowing them to rebind MHC molecules in the absence of stronger competing foreign epitopes.50–52 Indeed there is evidence that HLA-DP does not load via HLA-DM and preferentially presents self-antigens.53 54 Most tumors do not express MHC-II unless induced to do so by IFNγ. In this study with B16 tumors expressing MHC-II under a IFNγ promoter as in human tumors show that there is sufficient IFNγ produced to upregulate MHC-II. We thus propose a role for the direct recognition of tumor cells by CD4 T cells and suggest that naturally available IFNγ upregulates tumor MHC-II to facilitate this recognition and subsequent tumor clearance. It is likely that it is the CD4 T cells which are responsible for the production of the IFNγ required for MHC upregulation in the tumor environment through recognition of infiltrating APCs. Indeed we have demonstrated in our previous report that the anti-tumor effect mediated by the citrullinated vimentin peptides was reliant on the presence of IFNγ since blockade of IFNγ in vivo abrogated the tumor therapy.10
We have shown that a repertoire of T cells can be detected in healthy donors that are specific for the Modi-1 peptides. Citrullinated peptides can be presented in the thymus which may allow positive but not negative selection and result in a T cell repertoire recognizing these epitopes.55 There have been some studies showing a low frequency CD4 population in healthy donors to these citrullinated proteins.8 These studies were all performed in HLA-DR*0401 positive donors. In this study we have looked at donors with a variety of HLA types and have shown that although most responding donors/patients do express either HLA-DR*0401 or HLA-DP4 alleles at least one donor expressed neither allele. This suggests that responses to the Modi-1 peptides are able to be presented by another HLA allele in addition to HLA-DR4 and DP4 indicating a more promiscuous nature to these epitopes. This phenomenon has been previously reported in HIV infection where an HIV Gag peptide shows extensive MHC-II cross reactivity.56
In mouse models it has been shown that potentially pathogenic self-antigen-specific CD4 T cells are abundant in the repertoire of healthy mice and that these are kept under control by regulatory T cells.35 These studies show that vaccination overcomes this regulation resulting in potent anti-tumor immunity. In the human study we have depleted Tregs in order to assess the full functional T cell repertoire that could be expected following vaccination. We extended previous observations and have shown proliferative responses to one or more of the Modi-1 peptides in 71% healthy donors and 58% of ovarian cancer patients. No significant difference was observed between the proportion of responding T cells detected in healthy donors and ovarian cancer patients implying both possess a repertoire of T cells capable of responding to the Modi-1 peptides. The repertoires appear to show similar phenotypes in both healthy donors and the cancer patients although future characterization would benefit from determining if the responses exist in the naïve or memory pools.
We have shown that Modi-1 peptides when combined with a variety of TLR ligands stimulates high-frequency Th1 responses, and combining with TLR1/2 showed the strongest response in HLA-DR4 and DP4 transgenic mice. This highlights the importance of selecting the most appropriate TLR ligand for incorporating in a therapeutic peptide vaccine; this has also been demonstrated by others.57 58 In addition to the mixing of peptide vaccine with a TLR ligand, recent work has suggested that direct linkage of the peptide to the TLR ligand can dramatically enhance the immunogenicity of the vaccine.34 59–61 This enhancement is thought to be due to the direct targeting of the peptide and TLR ligand to the same APC, thus focusing the immune response. We next investigated if the direct linkage of the citrullinated peptides to the TLR1/2 ligand AMPLIVANT could induce a potent anti-tumor response. High frequency IFNγ responses were detected following the immunization of HLA-DR4 and DP4 transgenic mice with these linked peptides. Titration of the linked-peptide dose revealed that high frequency specific responses could be induced with at least a 10-fold lower peptide dose compared with the mixture of TLR ligand with peptide. This provides additional evidence that the linkage of a TLR ligand to a peptide enhances the immune responses and supports the development and clinical application of these peptide/TLR ligand linked conjugates.
Our study has shown the potential suitability of the Modi-1 vaccine for the treatment of ovarian cancers. We have shown that ovarian cancer patients have a repertoire of T cells that respond to the citrullinated vimentin and enolase peptides. The data presented in this study support the translation of the Modi-1 vaccine into the clinic for the treatment of patients with ovarian cancer.
Acknowledgments
Authors would like to thank Dr T Parsons, N Groves and Dr S Paston for critical reading of the manuscript.
References
Footnotes
Contributors VAB: Methodology, investigation, conceptualization, visualization, supervision, writing – original draft, review and editing; RLM: Conceptualization, methodology, resources; ID: Methodology, investigation; SA: Investigation; PS: Methodology, investigation; KWC: Methodology, investigation, visualization; MV: Formal analysis, writing – review and editing; RC: Investigation; PV: Investigation; MG: Methodology, investigation; GM: Resources; W-JK: Resources; CJM: Resources, writing – review and editing; LGD: Conceptualization, writing – original draft, writing – review and editing, supervision.
Funding This work was funded by Scancell Ltd, UK.
Competing interests VAB, RLM and LGD have ownership interest in patent WO2017013425 A1. LGD is the joint CEO of Scancell Ltd has ownership interest (including patents) in Scancell Ltd, is a consultant/advisory board member of Scancell Ltd, and has provided expert testimony for Scancell Ltd.
Patient consent for publication Not required.
Ethics approval All studies using human blood were approved by the local ethics committee and informed consent was obtained from all subjects involved. Samples were obtained from patients (ethics review: Nottingham BioBank APC170 no. 13-EM-0218 or IRAS project ID 238448) at Nottingham City Hospital and healthy donors (ethics review: 161–1711) from the University of Nottingham following University of Nottingham review and ethical approval. All animal work was carried out under a Home Office approved project license and in accordance with EU Directive 2010/63/EU.
Provenance and peer review Not commissioned; externally peer reviewed.
Data availability statement Data sharing not applicable as no data sets generated and/or analyzed for this study.