Article Text

Abstract
Background Antibody-drug conjugates are an exceptional and useful therapeutic tool for multiple diseases, particularly for cancer treatment. We previously showed that the fusion of the serine protease granzyme B (GrB), the effector molecule or T and B cells, to a binding domain allows the controlled and effective delivery of the cytotoxic payload into the target cell. The production of these constructs induced the formation of high molecular aggregates with a potential impact on the efficacy and safety of the protein.
Methods Our laboratory designed a new Fn14 targeted fusion construct designated GrB(C210A)-Fc-IT4 which contains a modified GrB payload for improved protein production and preserved biological activity. We assessed the construct’s enzymatic activity, as well as in vitro cytotoxicity and internalization into target cells. We also assessed pharmacokinetics, efficacy and toxicology parameters in vivo.
Results GrB(C210A)-Fc-IT4 protein exhibited high affinity and selective cytotoxicity within the nanomolar range when tested against a panel of Fn14-positive human cancer cell lines. The construct rapidly internalized into target cells, activating the caspase cascade and causing mitochondrial membrane depolarization. Pharmacokinetic studies in mice revealed that GrB(C210A)-Fc-IT4 displayed a bi-exponential clearance from plasma with a fast initial clearance (t1/2α=0.36 hour) followed by a prolonged terminal-phase plasma half-life (t1/2β=35 hours). Mice bearing MDA-MB-231 orthotopic tumor xenografts treated with vehicle or GrB(C210A)-Fc-IT4 construct (QODx5) demonstrated tumor regression and long-term (>80 days) suppression of tumor growth. Treatment of mice bearing established, subcutaneous A549 lung tumors showed impressive, long-term tumor suppression compared with a control group treated with vehicle alone. Administration of GrB(C210A)-Fc-IT4 (100 mg/kg total dose) was well-tolerated by mice and resulted in significant reduction of tumor burden in a lung cancer patient-derived xenograft model. Toxicity studies revealed no statistically significant changes in aspartate transferase, alanine transferase or lactate dehydrogenase in treated mice. Histopathological analysis of tissues from treated mice did not demonstrate any specific drug-related changes.
Conclusion GrB(C210A)-Fc-IT4 demonstrated excellent, specific cytotoxicity in vitro and impressive in vivo efficacy with no significant toxicity in normal murine models. These studies show GrB(C210A)-Fc-IT4 is an excellent candidate for further preclinical development.
- immunotherapy
- drug evaluation
- preclinical
- cytotoxicity
- immunologic
This is an open access article distributed in accordance with the Creative Commons Attribution Non Commercial (CC BY-NC 4.0) license, which permits others to distribute, remix, adapt, build upon this work non-commercially, and license their derivative works on different terms, provided the original work is properly cited, appropriate credit is given, any changes made indicated, and the use is non-commercial. See http://creativecommons.org/licenses/by-nc/4.0/.
Statistics from Altmetric.com
Background
Despite the numerous and successful advances in cancer therapeutic strategies in the past few years, there is an unmet need of developing drugs with completely new mechanisms of action. Monoclonal antibody (mAb) therapeutics have now become standard treatment for different disease indications including cancer, inflammatory and autoimmune diseases.1 More recently, antibody-drug conjugates (ADCs), which combine the cell binding specificity of mAbs with the cytotoxic potential of chemotherapeutic agents, have demonstrated significant clinical success. One of the most critical parameters in ADC design is selection of the antigen target because it has a direct impact on toxicity profile, efficacy and therapeutic window.2 3
The tumor necrosis factor (TNF) superfamily cytokine TNF-related weak inducer of apoptosis (TWEAK) acts via a cell surface receptor named Fn14, and the TWEAK/Fn14 axis is an important molecular pathway involved in the regulation of tissue responses following acute injury.4–8 Overexpression of Fn14 or TWEAK has been documented in many solid tumor types including lung, breast and prostate9–14 and has been shown to be an independent negative prognostic indicator in a variety of tumor types.6 14–21 Therefore, interruption of the TWEAK/Fn14 signaling axis or the use of the external receptor domain to deliver specific cytotoxic agents appears to be avenues for targeted therapy development.8 22
Previous studies have shown that anti-Fn14 antibodies are capable of suppressing the growth of tumor cells in vitro and in vivo.23–25 Clinical trials of antibodies targeting the TWEAK/Fn14 axis (drozitumab, enavatuzumab)26–28 have demonstrated safety and tolerability as well as antitumor effects in previously treated patients.
Our laboratory has previously reported that a human fusion protein construct targeting the Fn14 receptor and containing the cytotoxic granzyme B (GrB) payload (GrB-Fc-IT4) has in vitro and in vivo cytotoxic effects.7 During the course of our studies, however, we noticed the presence of high molecular aggregates during the production phase. We identified an unpaired Cys residue located in the C-terminal portion of GrB as a potential aggregation site. Here, we show the replacement of the impaired Cys had no impact on GrB enzymatic activity. In the present study we compared the in vitro cytotoxicity of GrB(C210A)-Fc-IT4 to GrB itself against a broad panel of non-small cell lung cancer (NSCLC) and other tumor cell lines. In addition, we conducted extensive mechanistic studies in vitro, as well as pharmacokinetic, toxicological and efficacy studies using several different in vivo tumor xenograft models.
Methods
Cell lines
MDA-MB-453, MDA-MB-468, MDA-MB-231, MEF 3.5−/−, A549, SKBR3, HT-29 and SKOV3 cell lines were obtained from the American Type Culture Collection and maintained as described previously described.29
Plasmid construction, protein expression and purification
To generate the GrB(C210A)-Fc-IT4, human GrB(C210A) cDNA was fused to the N-terminus of the coding region of hinge CH2 and CH3 (Fc) of a human IgG1 heavy chain followed by the humanized single-chain VH-VL variable fragment of the anti-Fn14 (scFvIT4) antibody using three separate PCR steps. The individual PCR fragments were then subjected to two splice overlap extension PCR reactions (first Fc and scFvIT4; then GrB(C210A) with Fc-IT4) to complete the fusion gene construct.30 The complete GrB(C210A)-Fc-IT4 fusion gene was then cloned into the mammalian expression vector pSecTag (Life Technologies, Waltham, Massachusetts, USA), expressed by transfection of HEK-293E cells and purified by immobilized metal affinity chromatography as previously described.7 The leader sequence containing the c-Myc epitope tag, (His)6 tag and the enterokinase cleavage site was cleaved by incubating the purified protein with recombinant enterokinase (New England Biolabs, Ipswich, Massachusetts, USA) overnight (20 U/mg) at room temperature (RT).31
GrB activity assay
GrB enzymatic activity was measured using a continuous colorimetric assay with Ac-IEPD-pNA (N-acetyl-Ile-Glu-Pro-Asp-p-nitroanilide) (Merck, Darmstadt, Germany) as a specific substrate as previously described.7 32
In vitro cytotoxicity assays
Cells were seeded onto plates overnight. After 24 hours incubation at 37°C, medium was replaced with complete media containing various concentrations of GrB or GrB(C210A)-Fc-IT4 and incubated at 37°C in 5% CO2 for 72 hours. Cell viability was determined using the crystal violet staining method as described previously.33
Internalization analysis
A549 and MDA-MB-231 cell lines were treated with 30 nmol/L GrB or GrB(C210A)-Fc-IT4 for various time periods. Internalization was assessed with an immunofluorescence assay as previously described.7 Studies to identify potential delivery of the GrB(C210A)-Fc-IT4 construct in the endosomal/lysosomal compartment were performed using the Lysotracker DND-22 kit (Life Technologies, Carlsbad, California, USA). A549 cells were treated with 30 nmol/L GrB(C210A)-Fc-IT4 labeled with Alexa Fluor 594 (Molecular Probes, Eugene, Oregon, USA) for 2 hours at 37°C, then media was replaced with fresh media containing the Lysotracker Blue DND-22 for 2 hours. Then cells were washed with phosphate buffered saline (PBS) and visualized with a Nikon A1Rsi multiphoton/confocal microscope using 405 nm and 561 nm lasers for excitation.
Apoptosis assays
Apoptosis was assessed using the Annexin V-FITC Kit (Invitrogen, Eugene, Oregon, USA).34 Mitochondrial membrane depolarization was studied using the cationic dye JC-1 (MitoProbe JC-1 Assay Kit, ThermoFisher Scientific, Eugene, Oregon, USA) as previously described.31
Western blot analysis
Western blot studies were performed using standard procedures.7 The primary antibodies used were anti-GrB (Santa Cruz Biotechnology, Santa Cruz, California, USA), anti-Caspase-3, anti-Caspase-7, anti-Caspase-9, anti-PARP-1 (Cell Signaling Technology, Danvers, Massachusetts, USA) and anti-β-actin (Sigma, St Louis, Missouri, USA).
Antibody dependent cell cytotoxicity assay
The antibody dependent cell cytotoxicity (ADCC) response was evaluated using the ADCC Reporter Bioassay kit (Promega, Madison, Wisconsin, USA). Briefly, SK-BR-3 cells were seeded onto plates and incubated overnight at 37°C. Serial dilutions of Trastuzumab (Roche, San Francisco, California, USA), an anti-CD20 antibody and GrB(C210A)-Fc-IT4 were prepared in RPMI-1640 media and added to SK-BR-3 cells. Jurkat/FcγRIIIa/NFAT-Luc cells were suspended in RPMI-1640 with low IgG serum and added to the cells, followed by incubation at 37°C. After 24 hours, luciferase activity was measured by ONE-Glo Luciferase Assay Reagent.
In vitro stability of GrB(C210A)-Fc-IT4
GrB(C210A)-Fc-IT4 (50 µg/mL) was incubated with commercial mouse serum (BioIVT, Westbury, New York, USA) for different time periods (0, 24, 48, 72, 96 hours) at 37°C. Samples were harvested on the same day and spun through microtainer vials. The anti-GrBhorseradish peroxidase (HRP) conjugated antibody (Santa Cruz Biotechnology) was used to detect the target protein and the signal intensity was quantitated by Adobe Photoshop with GrB(C210A)-Fc-IT4 as standard. The signal was normalized with a serum loading control (Transferrin, Genetex, Irvine, California, USA). Stability experiments were performed in duplicate.
Pharmacokinetic studies
BALB/c mice (Experimental Radiation Oncology Breeding Core, MD Anderson Cancer Center) at 4–6 weeks of age were injected (intravenous, tail vein) with 200 µg GrB(C210A)-Fc-IT4. At various time points (0–48 hours after fusion protein administration), mice were sacrificed and blood was extracted from the chest cavity. Plasma samples were analyzed using the Western blot method previously described as well as a quantitative ELISA assay. Briefly, ELISA microtiter plates (Santa Cruz Biotech) were coated (18 hours, 4°C) with 2 µg/mL of the Fn14 extracellular domain (Cell Signaling) in coating buffer (0.05 M Na2CO3, 0.05 M NaHCO3, pH 9.6) and blocked with 2% bovine serum albumin in PBS at RT for 2 hours. Samples (100 µL) were loaded in triplicate and incubated for 2 hours at RT, followed by the addition of HRP-conjugated goat anti-human IgG (KLP, Gaithersburg, Maryland, USA) in blocking buffer (1 hour, RT). Plates were washed five times after each step. The reaction was visualized by the addition of chromogenic substrate (ABTS) for 30 min and stopped with 10% sodium dodecyl sulfate (SDS). Absorbance at 450 nm was measured on a Thermomax plate reader. As a reference for quantification, a standard curve was established by a serial dilution of recombinant GrB(C210A)-Fc-IT4 in commercial mouse serum. Pharmacokinetic parameters were calculated by compartmental analysis of plasma data using an add-in program for pharmacokinetic and pharmacodynamic data analysis in Microsoft Excel the PKSolver (V.2.0).
In vivo efficacy using nude mice bearing human breast or lung cancer tumor xenografts
Groups of female BALB/c nude mice (MDACC Experimental Radiation Oncology Breeding Core) (7 weeks old, five mice per group) were injected with 5×106 human MDA-MD-231/Luc cells into the mammary fat pad (1:1 v/v with Matrigel). Once tumor volumes reached between 40 and 60 mm3, animals were randomized and treated (iv, tail vein) every other day for five total injections with vehicle or GrB(C210A)-Fc-IT4 (100 mg/kg total dose). Animals were monitored and tumor growth was measured every 3–4 days for an additional 90 days.
Human A549 lung cancer cells (7×106 cells) were subcutaneously injected on the right flank of 6-week-old female infant BALB/c nude mice. Once tumors were established (~25 mm3), animals were randomized and treated (intraperitoneal) every day for 10 total injections with vehicle, enavatuzumab (50 mg/kg) or GrB(C210A)-Fc-IT4 (50 mg/kg or 100 mg/kg total dose). Animals were monitored and tumor growth was measured every 3–4 days for 60 days.
In vivo efficacy study using lung cancer patient-derived xenograft model
Animal experiments were conducted following approval by the Institutional Animal Care and Use Committee guidelines of St. Joseph’s Hospital and Medical Center. Fresh lung tumors resected from a patient-derived xenograft (PDX) were subcutaneously (s.c.) implanted into the flank of 6-week-old nu/nu athymic mice (Jackson Labs). Tumors from the same patient xenografts were allowed to grow to approximately 200 mm3. Animals were grouped randomly (six mice per group) and treated with vehicle or GrB(C210A)-Fc-IT4 (20 mg/kg) every other day over a 10-day period for a total dose of 100 mg/kg via tail vein injection. Tumor size was measured 2–3 times per week for 30 days.
Toxicology studies
Female BALB/c mice (4–6 weeks old) were injected intravenously every other day for five total injections of vehicle or GrB(C210A)-Fc-IT4 (100 mg/kg total dose). One day after the last injection (Acute group, Day 10) or 4 weeks after the last injection (Recovery group, Day 38) animals in the study were euthanized and terminally bled for hematological and tissue harvest/histological analyzes. The studies were performed by the MDACC Department of Veterinary Medicine and Surgery.
Statistical analysis
All in vivo efficacy experiments, statistical analysis was performed using Prism V.8.0.0 (GraphPad Software). Comparisons of continuous variables between two groups were performed using unpaired t-test. Comparisons of continuous variables between three or more groups were performed using a one-way analysis of variance (ANOVA) followed by Dunnett’s multiple comparisons test. Differences were considered to be statistically significant when p values were <0.05.
Results
Construction, expression and purification of GrB(C210A)-Fc-IT4 fusion protein
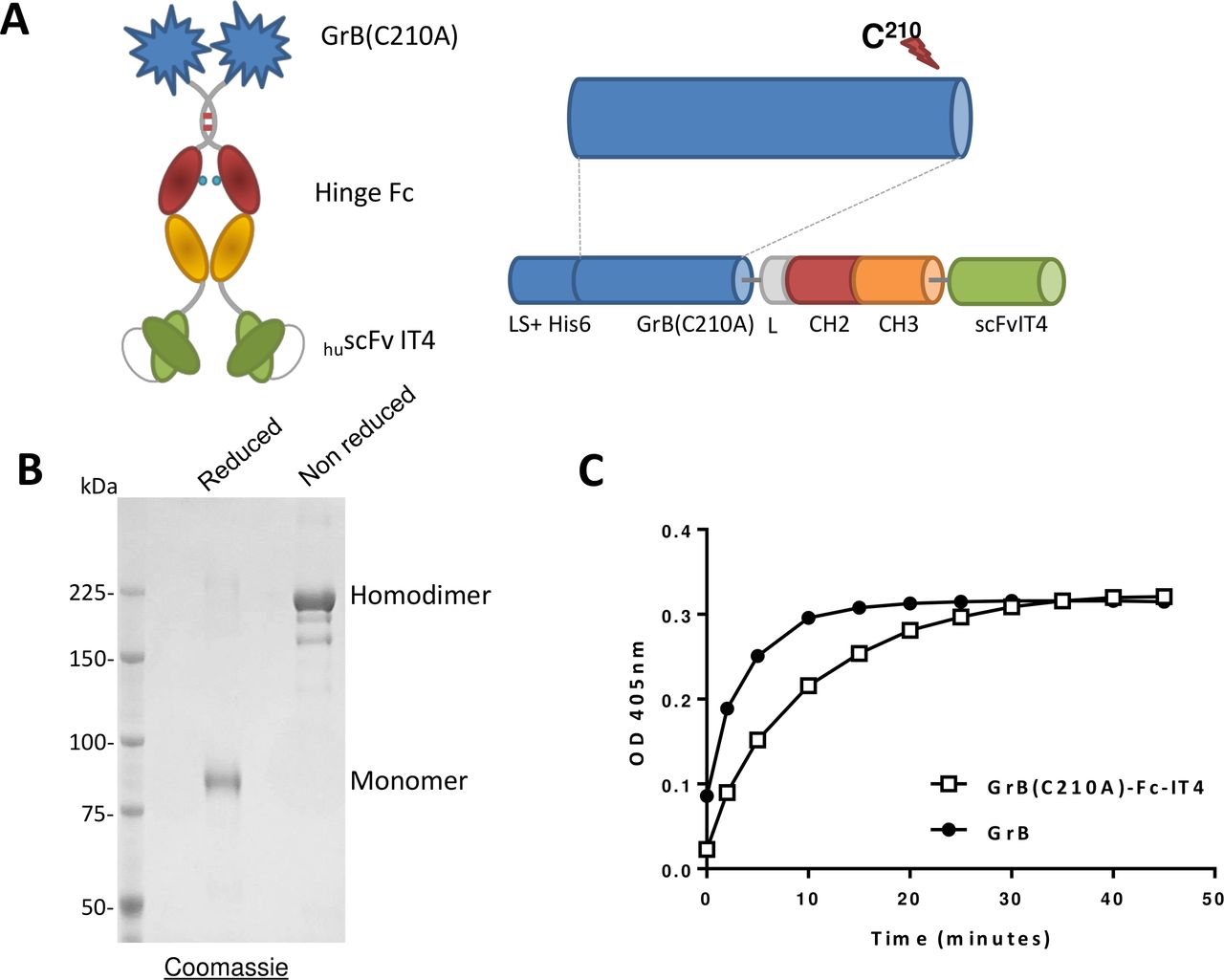
GrB(C210A)-Fc-IT4 (figure 1A) was transiently expressed by HEK-293E cells and purified to homogeneity. Analysis by SDS-PAGE of GrB(C210A)-Fc-IT4 under non-reducing conditions confirmed the purity of the construct (figure 1B). The theoretical molecular weight is 160 kDa for the homodimer but the presence of several glycosylation sites in the primary sequence increased the apparent molecular weight to 220 kDa. GrB(C210A)-Fc-IT4 had a similar enzymatic activity to commercial native GrB (264 U/nmol and 159 U/nmol, respectively), demonstrating the mutation has no impact on the enzymatic activity of GrB (figure 1C).
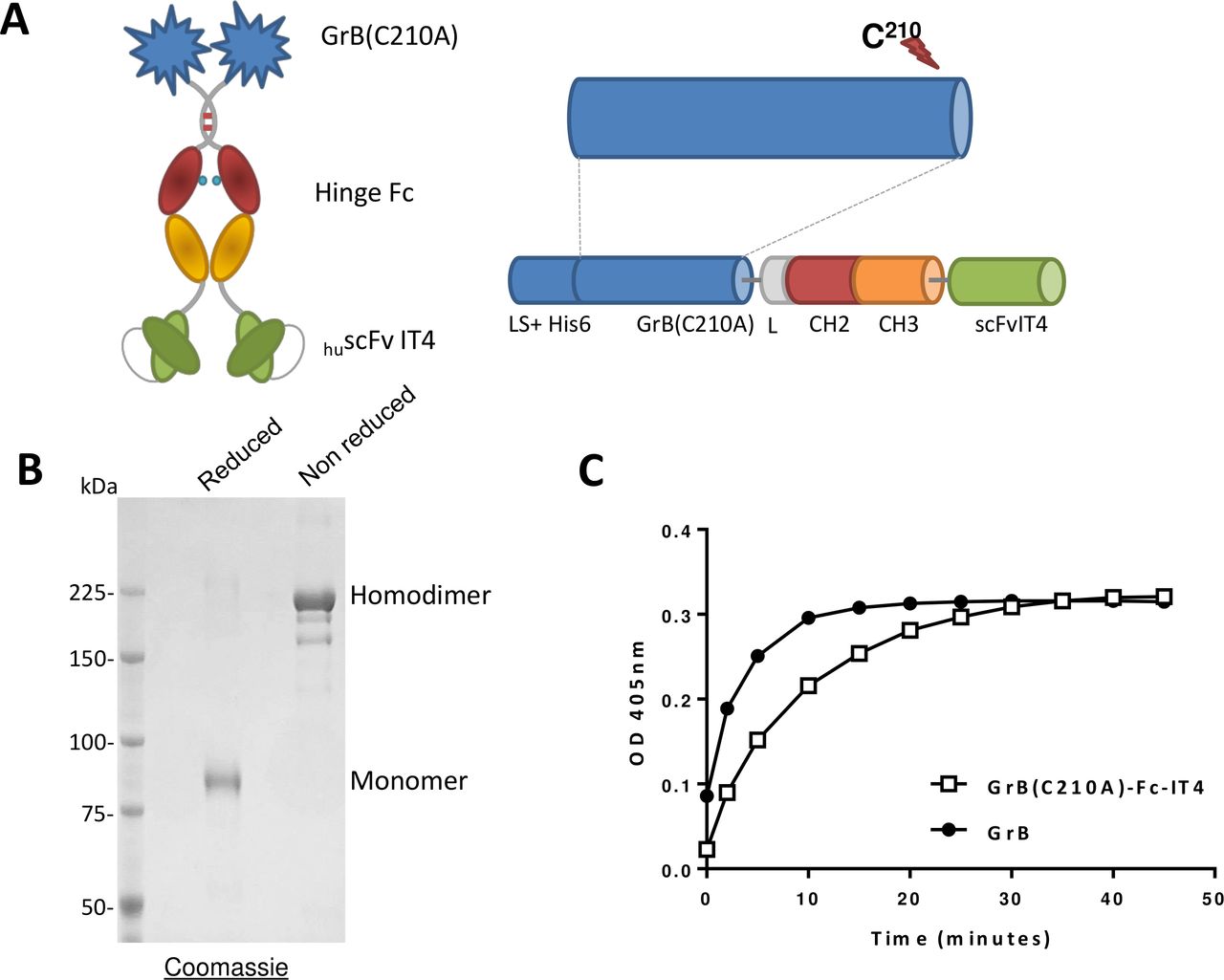
Schematic design, production and in vitro activity of the GrB(C210A)-Fc-IT4 protein. (A) Schematic diagram of the GrB(C210A)-Fc-IT4 construct. (B) SDS-PAGE analysis and Coomassie blue staining of purified GrB(C210A)-Fc-IT4 on a 6% gel run under reducing or non-reducing conditions. (C) GrB enzymatic activity. Purified GrB(C210A)-Fc-IT4 and GrB were incubated at 25°C with chromogenic substrate (Ac-IEPD-pNA) for the indicated times and absorbance was measured at 405 nm. His6, polyhistidine tag to facilitate purification; L-CH2-CH3, hinge and CH2 and CH3 domains of a human IgG1 heavy chain fragment; LS, the Igκ leader sequence that promotes the secretion of the fusion protein into the cell culture media; scFvIT4, humanized single chain fragment variable anti-Fn14 antibody.
Cytotoxic effect of GrB(C210A)-Fc-IT4 against various tumor cell lines and correlation with exposure time
Next, we assessed the cytotoxic effects of GrB(C210A)-Fc-IT4 on a panel of tumor cell lines (table 1). The IC50 of GrB(C210A)-Fc-IT4 ranged in the low nanomolar range, showing that the construct is able to induce cell death on Fn14 positive cell lines in a targeted manner. As previously published for the original GrB-Fc-IT4,7 we found no correlation between cell surface Fn14 levels and the sensitivity to the construct (data not shown). The fusion protein did not affect the growth of the mouse embryonic fibroblast MEF3.5−/− cell line, derived from Fn14 Knockout mice, indicating that the cytotoxicity of the construct on the tumor cell lines was dependent on the presence of the Fn14 receptor.
Comparative IC50 values of GrB and GrB(C210A)-Fc-IT4 against various cancer cell lines
We also examined the effect of GrB(C210A)-Fc-IT4 on MDA-MB-231 viability over time. While GrB(C210A)-Fc-IT4 treatment for 12 hours showed moderate levels of cytotoxicity, the lowest IC50 was obtained after 72 hours of continuous exposure (online supplemental table 1S). These results suggested cytotoxicity was due to receptor-mediated internalization and subsequent release of GrB(C210A)-Fc-IT4 into the cytosol.
Supplemental material
Internalization and intracellular localization of GrB(C210A)-Fc-IT4
To investigate the cytotoxic effect of the GrB(C210A)-Fc-IT4 on Fn14 positive cells, we first assessed the internalization process in MDA-MB-231 and A549 cells by immunofluorescence microscopy. We detected GrB(C210A)-Fc-IT4 in the cytoplasm within 30 min after exposure (figure 2A). In contrast, no staining was detected in cells treated with GrB alone, suggesting that internalization of GrB(C210A)-Fc-IT4 was driven by Fn14 receptor-mediated endocytosis. The intracellular delivery of the GrB portion and the metabolic fate of GrB(C210A)-Fc-IT4 was assessed at various times by Western blot analysis. Intact GrB(C210A)-Fc-IT4 was detected within the cell in less than 1 hour after exposure (figure 2B). Intracellular claeveage to release free GrB into the cytosol was observed at approximately 4 hours of treatment. Surprisingly, neither the intact GrB(C210A)-Fc-IT4 nor free GrB were observed after 16 hours, suggesting either complete degradation of the construct and its components or epitope masking of the GrB portion of the molecule.

Internalization and localization of GrB(C210A)-Fc-IT4. (A) MDA-MB-231 and A549 cells were treated with 30 nmol/L GrB or GrB(C210A)-Fc-IT4 for the indicated times. Cells were acid washed to remove surface bound material, fixed with phosphate buffered saline+3.7% paraformaldehyde, immunostained for the presence of GrB (green) and were counterstained with propidium iodide to identify nuclei (red). (B) Western blot analysis of the intracellular fate of 25 nmol/L GrB(C210A)-Fc-IT4 added to MDA-MB-231 cells positive controls are shown on the right side. (C) MDA-MB-231 cells were treated with different concentrations of GrB(C210A)-Fc-IT4 in the presence (25 µmol/L  ; 10 µmol/L
; 10 µmol/L ) or absence (
) or absence ( ) of the lysosomal inhibitor chloroquine and cell viability was measured at 72 hours. Cell viability was plotted as mean±SD of % of control. (D) Intracellular localization of GrB(C210A)-Fc-IT4. A549 cells were incubated for 2 hours with 30 nM Alexa-549-GrB(C210A)-Fc-IT4 (red) followed by addition of Lysotracker DND22 (blue). Cells were imaged live by confocal microscopy. Magnification 20×.
) of the lysosomal inhibitor chloroquine and cell viability was measured at 72 hours. Cell viability was plotted as mean±SD of % of control. (D) Intracellular localization of GrB(C210A)-Fc-IT4. A549 cells were incubated for 2 hours with 30 nM Alexa-549-GrB(C210A)-Fc-IT4 (red) followed by addition of Lysotracker DND22 (blue). Cells were imaged live by confocal microscopy. Magnification 20×.
Because accumulation of cell-targeted fusion proteins can occur in the endosomal and lysosomal compartments and can result in significant reduction in the efficiency of certain targeted therapeutics, we next evaluated the impact of chloroquine, a lysosomotropic agent that allows endosomal escape, on construct activity against MDA-MB-231 cells. Treatment of cells with different (non-cytotoxic) concentrations of chloroquine had no impact on the IC50 of GrB(C210A)-Fc-IT4 suggesting that endosomal or lysosomal trapping does not play a role in GrB(C210A)-Fc-IT4 internalization (figure 2C). We also studied the localization of GrB(C210A)-Fc-IT4 protein within the endosomes or lysosomes of A549 cells using Lysotracker Blue DND-22, a fluorescent dye that stains acidic compartments in live cells. We did not detect specific colocalization of the fusion protein in endosomes or lysosomes after 2 hours of treatment (figure 2D). Together, these studies confirm that GrB(C210A)-Fc-IT4 is internalized rapidly and efficiently escapes into the cytosol of target cells without appreciable accumulation in the lysosomal or endosomal compartments.
Mechanism of GrB(C210A)-Fc-IT4 induced apoptosis
GrB is a well-known potent initiator of apoptosis once delivered to the cytosol.35 To confirm that the cytotoxicity of GrB(C210A)-Fc-IT4 is mediated by apoptosis, we first assessed the Annexin V levels on MDA-MB-231 and A549 cell lines in the presence of different concentrations of either GrB(C210A)-Fc-IT4 or GrB. The construct GrB(C210A)-Fc-IT4 was able to increase Annexin V levels whereas GrB treatment alone showed no impact over 24 hours compared with non-treated cells (figure 3A). Similarly, treatment with GrB(C210A)-Fc-IT4 rapidly induced mitochondrial depolarization in treated cells over 24 hours (figure 3B).
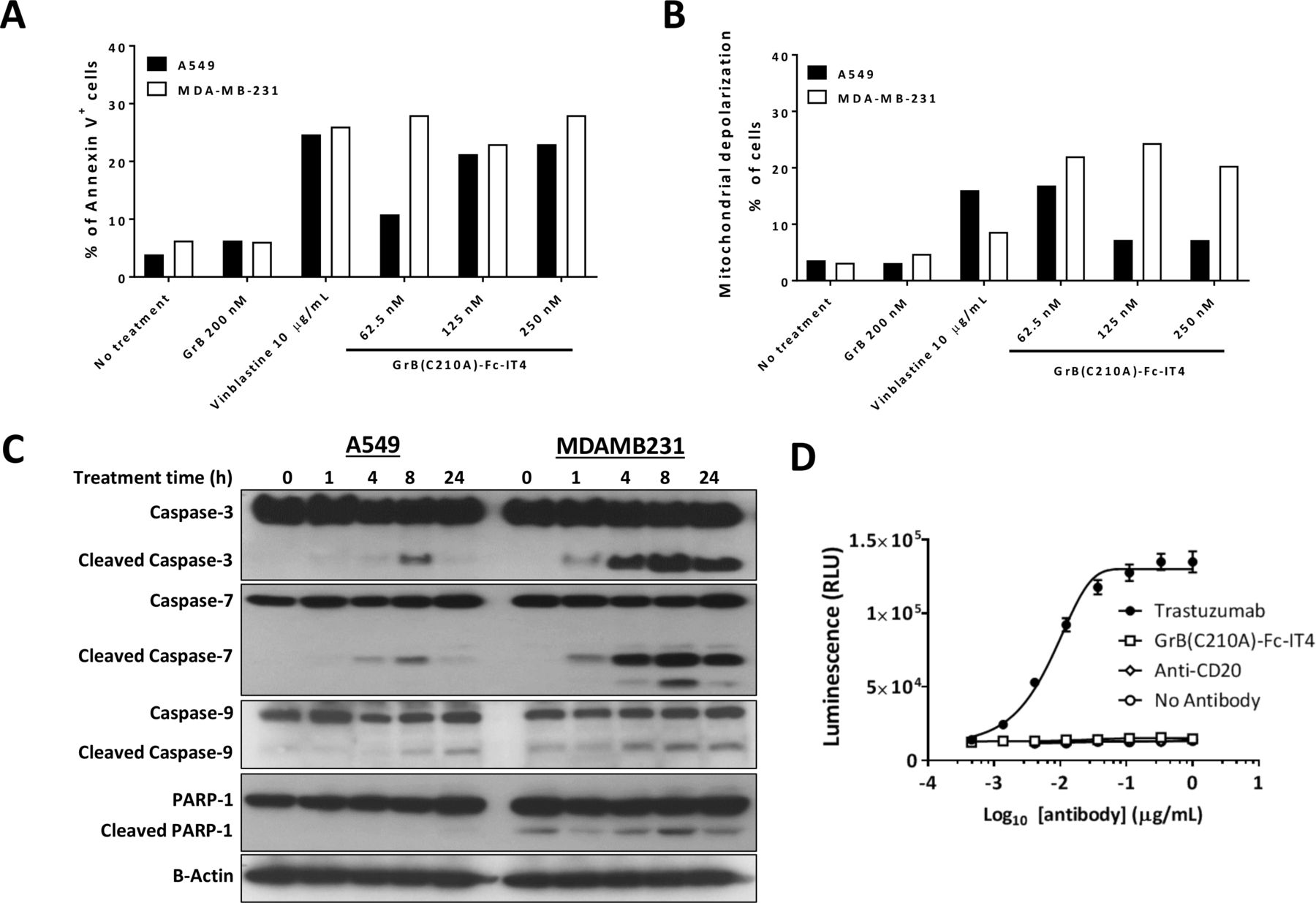
Mechanism of action of GrB(C210A)-Fc-IT4 fusion construct. (A) MDA-MB-231 and A549 cells were either left untreated or treated with the indicated agents at the indicated concentrations for 48 hours. Cells were then stained with Alexa Fluor 488 Annexin V and propidium iodide, followed by flow cytometric analysis. The percentage of annexin V+ cells includes early (Annexin V+ PI−) and late (Annexin V+ PI+) apoptosis. Vinblastine was used as a positive control for apoptosis. (B) MDA-MB-231 and A549 cells were either left untreated or treated and then assayed for mitochondrial membrane depolarization by flow cytometry. (C) MDA-MB-231 and A549 cells were left untreated or treated with 100 nmol/L GrB(C210A)-Fc-IT4 for the indicated times. Cells were collected and whole-lysates were analyzed by Western blot with the indicated antibodies. (D) SK-BR-3 cells were either left untreated or treated with GrB(C210A)-Fc-IT4, trastuzumab or a negative control antibody anti-CD20 at different concentrations. Antibody dependent cell cytotoxicity bioassay effector cells were added and after 24 hours induction at 37°C, Bio-Glo luciferase assay reagent was added and luminescence determined using a GloMax-Multi+Luminometer. The data were plotted as mean±SD and fitted to a 4PL curve using GraphPad Prism software.
Next, we used Western blot analysis to examine various cellular mediators of apoptosis known to be part of the GrB-induced apoptotic mechanism. Cells treated with GrB(C210A)-Fc-IT4 showed activation of the apoptotic cascade by cleavage of caspase-3, caspase-7, caspase-9 and PARP-1 at 8 hours after treatment (figure 3C). These data confirm that the mechanism of cytotoxic action appears to be similar to that of GrB itself.
An important mechanism of cytotoxic action of some conventional IgG surface-bound antibodies is the mediation of ADCC. To evaluate whether the Fc portion of the GrB(C210A)-Fc-IT4 is able to mediate ADCC on target cells, we performed an in vitro assay using the SK-BR-3 cell line which is known to express Fn14 and HER2 on the cell surface.36 As shown in figure 3D, treatment with the anti-HER2 antibody, trastuzumab, induced the activation of Jurkat cells whereas treatment with GrB(C210A)-Fc-IT4 and the irrelevant control antibody did not induce any response in the effector cells. This suggests that GrB(C210A)-Fc-IT4 is unable to induce ADCC likely due to the steric hindrance at the C-terminus of the Fc domain by the presence of the anti-Fn14 scFv domain. Therefore, all of the cytotoxic effects observed in vivo are likely to be solely due to the GrB component of the fusion construct.
Ex vivo stability and pharmacokinetics of GrB(C210A)-Fc-IT4
In order to assess the stability of the GrB(C210A)-Fc-IT4 molecule in a physiological environment, we incubated GrB(C210A)-Fc-IT4 in commercial mouse serum for up to 96 hours at 37°C and conducted Western blot analysis. Intact or full length GrB(C210A)-Fc-IT4 was present in serum for up to 96 hours with an overall ex vivo half-life of 88 hours, indicating the construct is stable under physiological conditions (figure 4A). In addition, quantitative ELISA and Western blot analysis of GrB(C210A)-Fc-IT4 from plasma samples harvested at 24, 48 and 72 hours after administration to BALB/c mice showed similar quantitative results (data not shown).

Stability, pharmacokinetics and toxicology of GrB(C210A)-Fc-IT4 in mice. (A) GrB(C210A)-Fc-IT4 was incubated in mouse serum (50 µg/mL) at 37°C for various lengths of time and the presence of intact GrB(C210A)-Fc-IT4 was assessed by Western blot analysis. Estimated percentage of the initial material was plotted as mean±SD versus time to obtain the ex vivo half-life. (B) GrB(C210A)-Fc-IT4 was injected via tail vein in BALB/c mice. Groups of mice (three mice per group) were sacrifice 0, 5, 15, 20 min, 1, 2, 4, 8, 16, 24 and 48 hours after injection. Heparinized blood samples were harvested from the chest cavity and centrifuged, the plasma fraction was analyzed for the presence of fusion protein using a GrB capture ELISA method. Pharmacokinetic parameters were calculated by compartmental analysis of plasma data using the PKSolver add-in program (V.2.0). t1/2α and t1/2β represent half-lives in the initial distribution phase and terminal elimination phase, respectively; (C) Body weight in vehicle group (phosphate buffered saline 10% Maltose) and GrB(C210A)-Fc-IT4 (100 mg/kg) treated mice. The values represent the weights measured in BALB/c mice before treatment, one after treatment (acute group) and 4 weeks after treatment (recovery group). (D) Liver enzymatic activity detected in serum of BALB/c treated mice. The enzymatic activity of ALT, AST and LDH was measured in acute and recovery groups. No statistical difference was found between mice treated with vehicle or GrB(C210A)-Fc-IT4 neither in the acute or the recovery groups. AUC0-∞, area under the blood concentration versus time curve; C0, estimated initial drug concentration in the blood; Cl, total body clearance; MRT, mean resident time; Vd, volume of distribution of the central compartment; Vss, volume of distribution at steady state.
Pharmacokinetic analysis of GrB(C210A)-Fc-IT4 revealed a biphasic clearance profile (figure 4B). Following intravenous administration, GrB(C210A)-Fc-IT4 demonstrated a rapid α-phase half-life of 0.36 hour, followed by a prolonged terminal β-phase clearance of 35.6 hours. The apparent small volume of distribution of 1.66 mL suggests limited distribution of GrB(C210A)-Fc-IT4 into peripheral sites outside the vascular space immediately after administration. The estimated initial (t=0) concentration in plasma was 120 µg/mL, similar to the projected initial dose concentration of 153 µg/mL of drug injected per mouse. These studies further confirm the stability of the molecule in vivo.
Toxicity studies of GrB(C210A)-Fc-IT4 in BALB/c mice
Next, we examined the in vivo toxicity observed in mice treated with total dose of 100 mg/kg of GrB(C210A)-Fc-IT4 using a QODX5 schedule. Body weight, tissue and serum samples were collected 24 hours (acute group) or 4 weeks (recovery group) after intravenous administration of the construct to BALB/c mice. There were no differences in body weight (figure 4C) or specific organ weight (data not shown) between the controls and the treated mice. The enzymatic activities of liver enzymes were determined and no differences in aminotransferase (AST), alanine transferase (ALT) and lactate dehydrogenase (LDH) levels between groups were found, indicating no evidence of hepatotoxicity (figure 4D). We performed a complete hematological and chemical analysis; we did not find significant differences between groups (data not shown). The histological analysis showed no moderate or marked lesions in any mice, and only a few rare and minor mild incidental lesions were present (online supplemental table 2S). Thus, GrB(C210A)-Fc-IT4 is a safe and well-tolerated drug at doses as high as 100 mg/kg in mice as there were no indications of severe toxicity.
In vivo efficacy of GrB(C210A)-Fc-IT4 in xenograft models
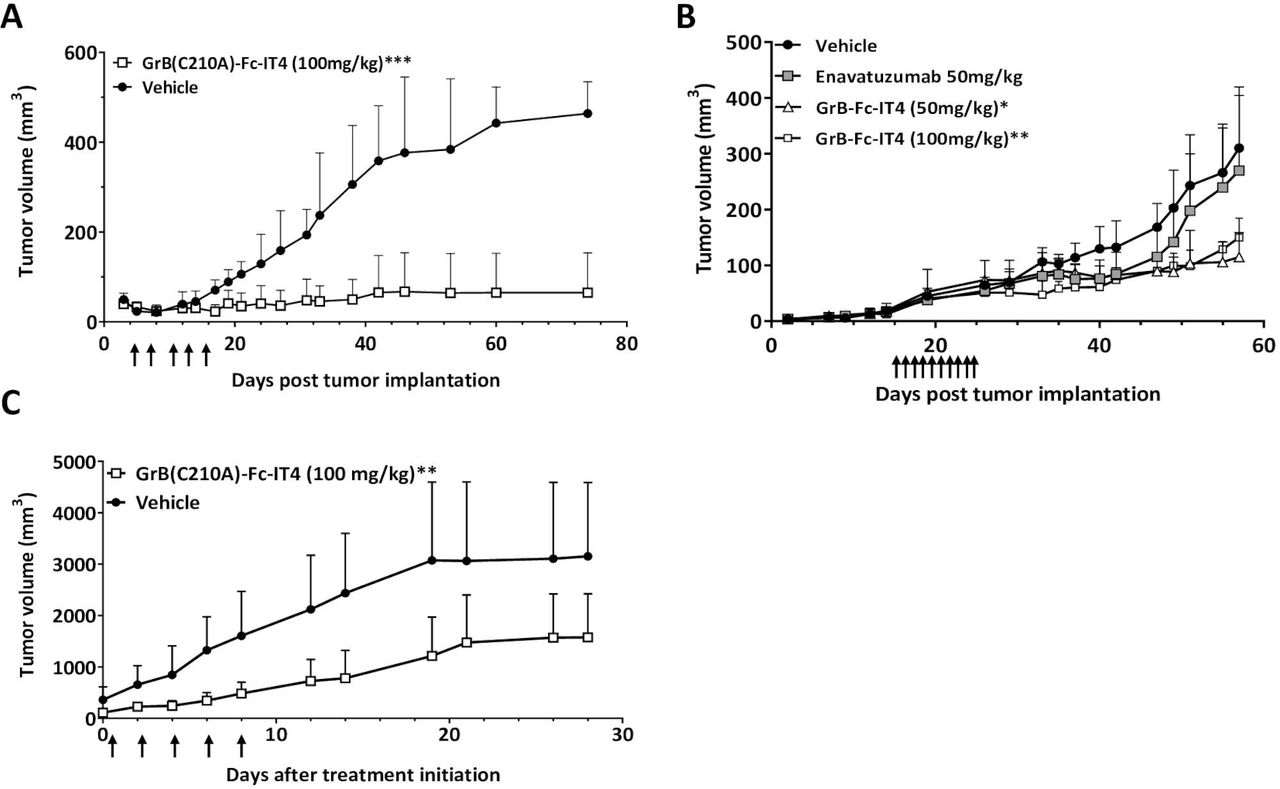
Once the pharmacokinetics and toxicity of GrB(C210A)-Fc-IT4 were determined, we then evaluated the ability of GrB(C210A)-Fc-IT4 to inhibit the growth of established MDA-MB-231 tumors in BALB/c nude mice. Three out of five mice with tumors treated every other day with GrB(C210A)-Fc-IT4 for 10 days showed complete tumor growth inhibition whereas the other two mice showed a significant tumor growth inhibition (p<0.001) compared with control (vehicle) tumors 80 days after implantation (figure 5A). Furthermore, mice bearing MDA-MB-231 tumors treated via intravenous or intraperitoneal injection with GrB(C210A)-Fc-IT4 (100 mg/kg) following the same scheduled (QODX5) showed significant tumor growth delay independent of the route of administration (online supplemental figure 1S). There were no changes in body weight in control versus treated mice for the duration of both studies indicating that 100 mg/kg total dose was well tolerated in mice.
Supplemental material

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
In vivo activity of the GrB(C210A)-Fc-IT4 fusion construct on different tumor models. (A) MDA-MB-231-Luc cells were implanted under the mammary fat pad of nude mice and groups of mice (n=5) were treated intravenous with vehicle (phosphate buffered saline (PBS) 10% maltose ) or GrB(C210A)-Fc-IT4 (100 mg/kg
) or GrB(C210A)-Fc-IT4 (100 mg/kg ). Treatment was every other day when the tumors reached approximately 75 mm3, arrows represent treatment day. Tumor size was assessed by direct caliper measurement. Efficacy data were plotted as mean tumor volume (in mm3)±SD. ***p<0.0003, compared with vehicle group. (B) A549 cells were implanted subcutaneously on the right flank of nude mice and groups of mice (n=5) were treated intraperitoneal with vehicle (PBS 10% maltose
). Treatment was every other day when the tumors reached approximately 75 mm3, arrows represent treatment day. Tumor size was assessed by direct caliper measurement. Efficacy data were plotted as mean tumor volume (in mm3)±SD. ***p<0.0003, compared with vehicle group. (B) A549 cells were implanted subcutaneously on the right flank of nude mice and groups of mice (n=5) were treated intraperitoneal with vehicle (PBS 10% maltose ), enavatuzumab (50 mg/kg
), enavatuzumab (50 mg/kg ) or GrB(C210A)-Fc-IT4 (50 mg/kg
) or GrB(C210A)-Fc-IT4 (50 mg/kg  , 100 mg/kg
, 100 mg/kg ). Treatment was every day (10 doses) when the tumors reached approximately 25 mm3. Arrows represent treatment day. Tumor size was assessed by direct caliper measurement. Efficacy data were plotted as mean tumor volume (in mm3)±SD. *p<0.0382, **p<0.0337, compared with vehicle group. (C) Fresh tumor cells from a patient were implanted subcutaneously (s.c.) into the flanks of 6-week-old nude mice, when the tumors reached approximately 200 mm3 animals were grouped randomly (n=6) and treated every other day intravenous route with vehicle (PBS
). Treatment was every day (10 doses) when the tumors reached approximately 25 mm3. Arrows represent treatment day. Tumor size was assessed by direct caliper measurement. Efficacy data were plotted as mean tumor volume (in mm3)±SD. *p<0.0382, **p<0.0337, compared with vehicle group. (C) Fresh tumor cells from a patient were implanted subcutaneously (s.c.) into the flanks of 6-week-old nude mice, when the tumors reached approximately 200 mm3 animals were grouped randomly (n=6) and treated every other day intravenous route with vehicle (PBS ), or GrB(C210A)-Fc-IT4 (100 mg/kg
), or GrB(C210A)-Fc-IT4 (100 mg/kg ). **p<0.0042, compared with vehicle. Efficacy data were plotted as mean tumor volume (in mm3)±SD. Treatment days are indicated with arrows.
). **p<0.0042, compared with vehicle. Efficacy data were plotted as mean tumor volume (in mm3)±SD. Treatment days are indicated with arrows.
In order to determine whether the antitumor effect observed was mediated by the cytotoxic payload (GrB) and not because of TWEAK/Fn14 axis inhibition by the scFvIT4 targeting moiety, we treated nude mice with subcutaneous A549 NSCLC tumors every day for 10 days with either GrB(C210A)-Fc-IT4 or the unconjugated anti-Fn14 antibody, enavatuzumab at equimolar drug concentrations.27 28 Administration of enavatuzumab resulted in transient tumor growth inhibition followed by rapid regrowth of the tumors back to and exceeding control tumor growth (figure 5B). In contrast, treatment with our GrB-based construct was able to induce a sustained prolonged growth delayed when compared with controls.
Finally, we examined the efficacy of GrB(C210A)-Fc-IT4 in a human lung PDX model. Tumor cells were subcutaneously (s.c.) implanted in the flank of mice and once tumors reached about 200 mm3, animals were grouped randomly (six mice per group) and treated with PBS or GrB(C210A)-Fc-IT4 (100 mg/kg). We observed a 50% reduction in tumor burden in GrB(C210A)-Fc-IT4 treated mice compared with the vehicle group (figure 5C). Overall, these data indicate that GrB(C210A)-Fc-IT4 is effective against breast and lung tumor xenograft models causing a dramatic and prolonged tumor growth inhibition, and also reduces tumor growth rate in a lung PDX tumor model.
Discussion
Cancer treatment has been evolving over the years, from basic strategies including surgery, radiation and systemic chemotherapy to more elaborate targeted approaches such as immunotherapy and personalized molecularly targeted medicine. In recent years, the use of chemotherapy has been augmented thanks to the development of targeted therapies like kinase inhibitors, ADCs or similar molecules.37 We have previously demonstrated that the GrB-based fusion construct targeting Fn14 is an effective and powerful therapeutic.7 In this study we extended these findings and demonstrated that the specific mutation introduced into the new GrB(C210A)-Fc-IT4 fusion construct did not impact the potent cytotoxic effect of the drug against a large panel of tumors cells. GrB(C210A)-Fc-IT4 is able to internalize into Fn14 positive cells in a rapid and specific manner via endocytosis and once it is in the cytoplasm it induces cell death via the GrB portion of the construct.
GrB is a highly cytotoxic serine protease expressed by human immune effector cells and is the causative agent whereby these cells induce pro-apoptotic signals when delivered to the cytoplasm of target cells.38 The protein has been well studied and is known to operate through multimodal cascade pathways, which are both caspase dependent and independent. We have previously shown that GrB-based fusion constructs mechanism of action includes activation of the caspase cascade and release of cytochrome c from the mitochondrial compartment7 resulting in activation of a caspase-independent apoptotic cascade exemplified by mitochondrial depolarization. The well-known trimodal mechanism of GrB cytotoxic activity is different from that of the small cytotoxic molecules used in other ADCs such as auristatin or maytansine derivatives which are microtubule inhibitors.39 Importantly, while intrinsic resistance mechanisms such as multidrug resistance (MDR) drug efflux pumps are known to confer resistance to apoptosis, previous studies showed that expression of MDR did not result in resistance mechanisms to GrB-based targeted fusion constructs like GrB(C210A)-Fc-IT4.7
The cell surface receptor Fn14 for the cytokine TWEAK has emerged as a valuable target for cancer therapy because it displays low expression in most normal tissues and has been shown to be significantly elevated in a variety of solid tumor types.7 9–13 40 41 Previously we were not able to find a correlation between the level of Fn14 overexpression in cancer cells and sensitivity to GrB(C210A)-Fc-IT4. We have shown similar results with other GrB-based constructs targeting different molecules like HER2 or VEGFR2.7 31 42 These data suggest that target abundance alone does not determine sensitivity. Other mechanisms such as target availability, endocytosis, endosomal release or apoptosis induction by GrB may play a determining factor. We are currently performing additional studies to explore more closely the intracellular elements related to the signaling pathways involved in the GrB cytotoxic mechanism(s) of action.
An important parameter in the development of antibody drug conjugates is pharmacokinetics studies to provide critical information about drug’s behavior and metabolism in vivo. In our ex vivo studies we were able to demonstrate intact GrB(C210A)-Fc-IT4 up to 96 hours in circulation. We were unable to analyze for specific GrB enzymatic activity, because endogenous levels of free GrB present in the serum would interfere with the assay. The pharmacokinetic analysis showed that GrB(C210A)-Fc-IT4 has a secondary half-life over 35 hours in BALB/c mice, with a plasma concentration over 10 µg/mL at 48 hours after injection. Although GrB(C210A)-Fc-IT4 has a fast initial clearance, it is highly stable in circulation, which corresponds with bi-exponential kinetics. As expected, in our design of the construct, the dimeric molecular weight of GrB(C210A)-Fc-IT4 offered a prolonged circulation in blood of mice, which potentially allows higher accumulation within a tumor. It is important to note that the original anti-Fn14 antibody we employed to generate the IT4 scFv (ITEM4) cross-reacts with the murine Fn14 receptor.43 Therefore, toxicity in normal murine tissues observed with GrB(C210A)-Fc-IT4 will be important indicators to consider for human clinical trials. In addition, other studies have shown that human GrB is approximately 30-fold more toxic than murine GrB.44 Therefore, toxicity (if any) observed by the GrB(C210A)-Fc-IT4 construct studies in mice may over predict for clinical toxicity. Our toxicity studies performed in BALB/c mice without tumors showed that GrB(C210A)-Fc-IT4 is a safe and tolerable drug at doses as high as 100 mg/kg. Moreover, we did not find signs of hepatotoxicity in the clinical chemistry or the histological analyzes. This is a compelling result as compared with other anti-Fn14 antibodies, like enavatuzumab, which induced mild elevations of the liver enzymes (ALT, AST) in their preclinical studies in non-human primates at the same dose.28 Because of the absence of toxicity at the doses tested, we have yet to establish a maximum tolerated dose (MTD) for GrB(C210A)-Fc-IT4. Studies suggest that the MTD is likely above the efficacious 100 mg/kg total dose level employed in our studies.
The efficacy studies against MDA-MB-231 and A549 tumor xenografts and lung PDX tumor models demonstrated excellent in vivo tumor growth suppression. In addition, no significant difference was observed in the efficacy of mice treated via intravenous or intraperitoneal injection. The in vivo efficacy comparison between GrB(C210A)-Fc-IT4 and enavatuzumab in the A549 xenograft tumor model demonstrated that GrB plays a critical role in mediating the cytotoxicity with a negligible role played by the targeting moiety. All these results together demonstrate that GrB(C210A)-Fc-IT4 is an effective antitumor agent that should be considered for clinical evaluation in patients with Fn14-positive tumors.
Supplemental material
References
Supplementary materials
Supplementary Data
This web only file has been produced by the BMJ Publishing Group from an electronic file supplied by the author(s) and has not been edited for content.
Footnotes
Correction notice This article has been corrected since it was published. The author name ‘Walter N Hittelman’ was incorrectly spelt as ‘Walter N Hittlerman’.
Contributors Conception and study design: AAdC, LHC, KAM and MGR. Development of methodology: AAdC, LHC, TGW, JAW, WNH, KAM and MGR. Data acquisition: AAdC, LHC, TGW, JAW, WNH, KAM and MGR. Analysis and interpretation of data: AAdC, LHC, TGW, WNH, KAM and MGR. Manuscript preparation: AAdC, LHC, WNH, KAM, JAW, TGW and MGR. Study supervision: AAdC and MGR. All authors read and approved the final manuscript.
Funding Research supported by NCI # CA16672 (M.D. Anderson Cancer Center). Research conducted, in part, by the Clayton Foundation for Research. Flow cytometry experiments performed by the Flow cytometry and cellular imaging core facility: this shared resource is partially funded by NCI Cancer Center Support Grant P30CA16672.
Competing interests AAdC, LHC, KAM and MGR report grants from the Clayton Foundation for Research and Mirata Biopharma, during the conduct of the study. In addition, MGR, KAM and LHC have a patent on Serine protease molecules and therapies issued.
Patient consent for publication Not required.
Ethics approval All mice were maintained under specific pathogen–free conditions according to the American Association for Accreditation of Laboratory Animal Care (AAALAC) standards. All experimental research on mice was done in accordance with protocols approved by MD Anderson Cancer Center.
Provenance and peer review Not commissioned; externally peer reviewed.
Data availability statement All data relevant to the study are included in the article or uploaded as supplementary information. Data are available upon reasonable request.