Article Text

Abstract
Combined radioimmunotherapy is currently being investigated to treat patients with cancer. Anti-programmed cell death-1 (PD-1) immunotherapy offers the prospect of long-term disease control in solid tumors. Radiotherapy has the ability to promote immunogenic cell death leading to the release of tumor antigens, increasing infiltration and activation of T cells. New York esophageal squamous cell carcinoma-1 (NY-ESO-1) is a cancer–testis antigen expressed in 20% of advanced gastric cancers and known to induce humoral and cellular immune responses in patients with cancer. We report on the dynamic immune response to the NY-ESO-1 antigen and important immune-related biomarkers in a patient with metastatic gastric cancer treated with radiotherapy combined with anti-PD-1 pembrolizumab antibody.Our patient was an 81-year-old man diagnosed with locally advanced unresectable mismatch repair-deficient gastric cancer having progressed to a metastatic state under a second line of systemic treatment consisting of an anti-PD-1 pembrolizumab antibody. The patient was subsequently treated with local radiotherapy administered concomitantly with anti-PD-1, with a complete response on follow-up radiologic assessment. Disease control was sustained with no further therapy for a period of 12 months before relapse. We have identified an NY-ESO-1-specific interferon-γ (IFN-γ) secretion from the patients’ T cells that was significantly increased at response (****p˂0.0001). A novel promiscuous immunogenic NY-ESO-1 peptide P39 (P153–167) restricted to the four patient’s HLA-DQ and HLA-DP alleles was identified. Interestingly, this peptide contained the known NY-ESO-1-derived HLA-A2-02:01(P157–165) immunogenic epitope. We have also identified a CD107+ cytotoxic T cell subset within a specific CD8+/HLA-A2-NY-ESO-1 T cell population that was low at disease progression, markedly increased at disease resolution and significantly decreased again at disease re-progression. Finally, we identified two groups of cytokines/chemokines. Group 1 contains five cytokines (IFN-γ, tumor necrosis factor-α, interleukin-2 (IL-2), IL-5 and IL-6) that were present at disease progression, significantly downregulated at disease resolution and dramatically upregulated again at disease re-progression. Group 2 contains four biomarkers (perforin, soluble FAS, macrophage inflammatory protein-3α and C-X-C motif chemokine 11/Interferon–inducible T Cell Alpha Chemoattractant that were present at disease progression, significantly upregulated at disease resolution and dramatically downregulated again at disease re-progression. Combined radioimmunotherapy can enhance specific T cell responses to the NY-ESO-1 antigen that correlates with beneficial clinical outcome of the patient.
- antigens
- neoplasm
- biomarkers
- tumor
- case reports
- CD4-CD8 ratio
- combined modality therapy
This is an open access article distributed in accordance with the Creative Commons Attribution Non Commercial (CC BY-NC 4.0) license, which permits others to distribute, remix, adapt, build upon this work non-commercially, and license their derivative works on different terms, provided the original work is properly cited, appropriate credit is given, any changes made indicated, and the use is non-commercial. See http://creativecommons.org/licenses/by-nc/4.0/.
Statistics from Altmetric.com
Background
With 1 033 701 new cases reported worldwide in 2018, gastric cancer ranks as the fifth cancer in incidence and third in mortality according to the latest Globocan estimates.1 Standard of care treatment of unresectable disease has long been restricted to palliative chemotherapy offering modest benefit, with 5-year survival rates inferior to 12 months. More recently, the development of immune checkpoint inhibitors (ICIs) targeting the programmed cell death-1 (PD-1)/programmed cell death ligand-1 (PD-L1) pathway has led to substantial therapeutic improvements in several solid malignancies. However, the benefit of these novel therapies in gastric cancer has been restricted to a small minority of patients, with reported objective response rates ranging from 12% to 25%.2 Higher response rates are however reported for tumors displaying microsatellite instability (MSI-H) with phenotypic evidence of mismatch repair deficiency (MMR-deficient).3 MSI can be found in 22% of gastric cancers diagnosed in western countries and defines one of the four distinct gastric cancer subgroups.3 The high mutation rate inherent to the underlying genetic alterations of MSI-H tumors confers them with a high mutational load.4 A high tumor mutational burden (TMB), through an increased probability of immunogenic neoantigen expression, has been shown to predict response to anti-PD-1 ICIs in a given tumor type.5 The capacity of TMB to predict ICI treatment outcome is nonetheless inconsistent6 and despite showing comparatively high sensitivity to anti-PD-1, almost half of MSI-H tumors will not respond to these therapies.
One drawback of TMB is that despite correlating with the quantity of neoantigens expressed by a tumor, it fails to account for the intrinsic properties of the latter. It is now becoming apparent that some neoantigens are more immunogenic than others. The cancer–testis antigens (CTAs) represent a distinctive category of tumor antigens characterized by high cancer specificity and high immunogenicity.7 New York esophageal squamous cell carcinoma-1 (NY-ESO-1) is a CTA reported to be expressed in up to 20% of patients with metastatic gastric cancer,8 having the capacity to induce both natural antibody and T cell responses.9 Moreover, anti-NY-ESO-1 immunity has been demonstrated to be associated with an improved clinical benefit in ipilimumab (anti-cytotoxic T-lymphocyte-associated protein 4)-treated melanoma.10 We have also demonstrated recently that the presence of anti-NY-ESO-1-integrated immunity may potentially identify a response to PD-1 blockade in patients with head and neck squamous cell carcinoma.11
Recent developments point to the tumor microenvironment (TME) as another important determinant of anti-PD-1 efficacy. A high pretreatment concentration of tumor-infiltrating lymphocytes along with appropriate immunostimulatory conditions have been shown to be strongly associated with the probability of response to ICI.12 Radiotherapy (RT) has shown interesting properties in this regard, through its ability to promote immunogenic cell death leading to the release of tumor neoantigens, increasing infiltration and activation of T cells and consequently enhancing the T cell repertoire.13 This synergistic effect of RT on ICI efficacy has been observed in a number of preclinical models.14 Furthermore, RT provides a rational for a phenomenon known as the abscopal effect. In this, ICI-treated patients given RT at one tumor site present an immune-mediated response in distant non-irradiated lesions.15
In this study, we report on a patient with an MMR-deficient non-resectable gastric cancer who displayed a long-lasting response to an anti-PD-1 treatment triggered by a localized RT that correlated with a sustained NY-ESO-1-specific T cells response.
Case report
We report on an 81-year-old male patient diagnosed with an MMR-deficient HER-2 negative gastric adenocarcinoma staged cT4cN3M0 at disease presentation. He was offered upfront chemotherapy with four cycles of FOLFOX chemotherapy complicated by grade IV fatigue. The tumor was found to be unresectable on laparoscopic exploration, leading to abortion of the surgical procedure. Considering the relatively poor tolerance to the upfront chemotherapy, a second-line treatment with the anti-PD-1 ICI pembrolizumab was subsequently initiated (figure 1A) based on the MMR deficiency status of the tumor. The treatment was interrupted after the administration of the sixth cycle due to grade III liver toxicity. Following a 2-month treatment interruption marked by clinical stability and resolution of liver function tests, a radiologic assessment was repeated, revealing frank progression of the primary lesion (figure 1B–a) along with the development of new locoregional and retroperitoneal metastatic lymph nodes (progression stage). The patient was then complaining of an epigastric pain coinciding with a persistent low-grade bleeding from the primary tumor and was addressed for RT for hemostatic and locoregional control purposes. The patient received a volumetric intensity-modulated RT treatment on the primary tumor along with the enlarged abdominal lymph nodes for a total of 45 Gy in 1.8 Gy fractions (figure 1A). Of note, the pembrolizumab treatment was resumed along with the initiation of the RT for three additional doses before being permanently interrupted based on the patient’s wishes. The positron emission tomography-computed tomography (PET–CT) repeated 2 months after the term of the RT revealed a near-complete resolution (disease resolution stage) of the gastric uptake along with the disappearance of the locoregional and metastatic lymph nodes (figure 1B–b). Follow-up showed a subsequent long-lasting disease control of 12 months before a relapse marked by locoregional progression (re-progression stage) and diffuse distant metastatic spread (figure 1B–c). The clinical course of the disease, the different time-points for collection of blood samples and time-points for pembrolizumab cycles, RT sessions and PET/CT imaging are summarized in figure 1A,B.

Clinical course of disease and time-points for collection of blood samples, for pembrolizumab (Pembro) cycles, radiotherapy sessions and PET–CT imaging. (A) On the left: dates for pembrolizumab cycles and for PET–CT scan. On the middle (red shapes): dates for blood samples collection. On the right: radiotherapy duration. (B) PET–CT scan images showing the clinical evolution of the disease at different time-points before and after combined radiotherapy–pembrolizumab treatment. (a) PET–CT conducted in November 2017 showed disease progression with increased metabolism in the primary lesion and development of multiple new abdominal metastatic lymph nodes. (b) Restaging with PET–CT done in February 2018 showed a near-complete resolution of gastric wall uptake with complete disappearance of locoregional and other retroperitoneal lymph nodal activities. (c) PET–CT done on February 27, 2019, revealed disease re-progression with extent of intense uptake in the stomach. PET, positron emission tomography.
Determination of antitumor immune responses
Evaluation of anti-NY-ESO-1 antibody levels and characterization of anti-NY-ESO-1-specific T cell response
The anti-NY-ESO-1 antibody was measured by ELISA assays (see Materials and Methods in online supplemental file 1). We first determined an optical density (OD) cutoff value for the NY-ESO-1 humoral immune response positivity. In this, we used OD values from pooled plasma samples obtained from five healthy donors and two patients with cancer negative for NY-ESO-1 as the cutoff for NY-ESO-1 positivity at a dilution of 1:100. Plasma samples at different stages (disease progression, disease resolution and disease re-progression) were assayed. A weak positive reaction was exhibited in which the OD values slightly exceeded the cutoff, but no significant difference was observed between the three different disease stages (online supplementary figure S1). Similarly, no significant difference was recorded when the results were compared with data obtained with the pooled serum from five healthy donors used as negative control.
Supplemental material
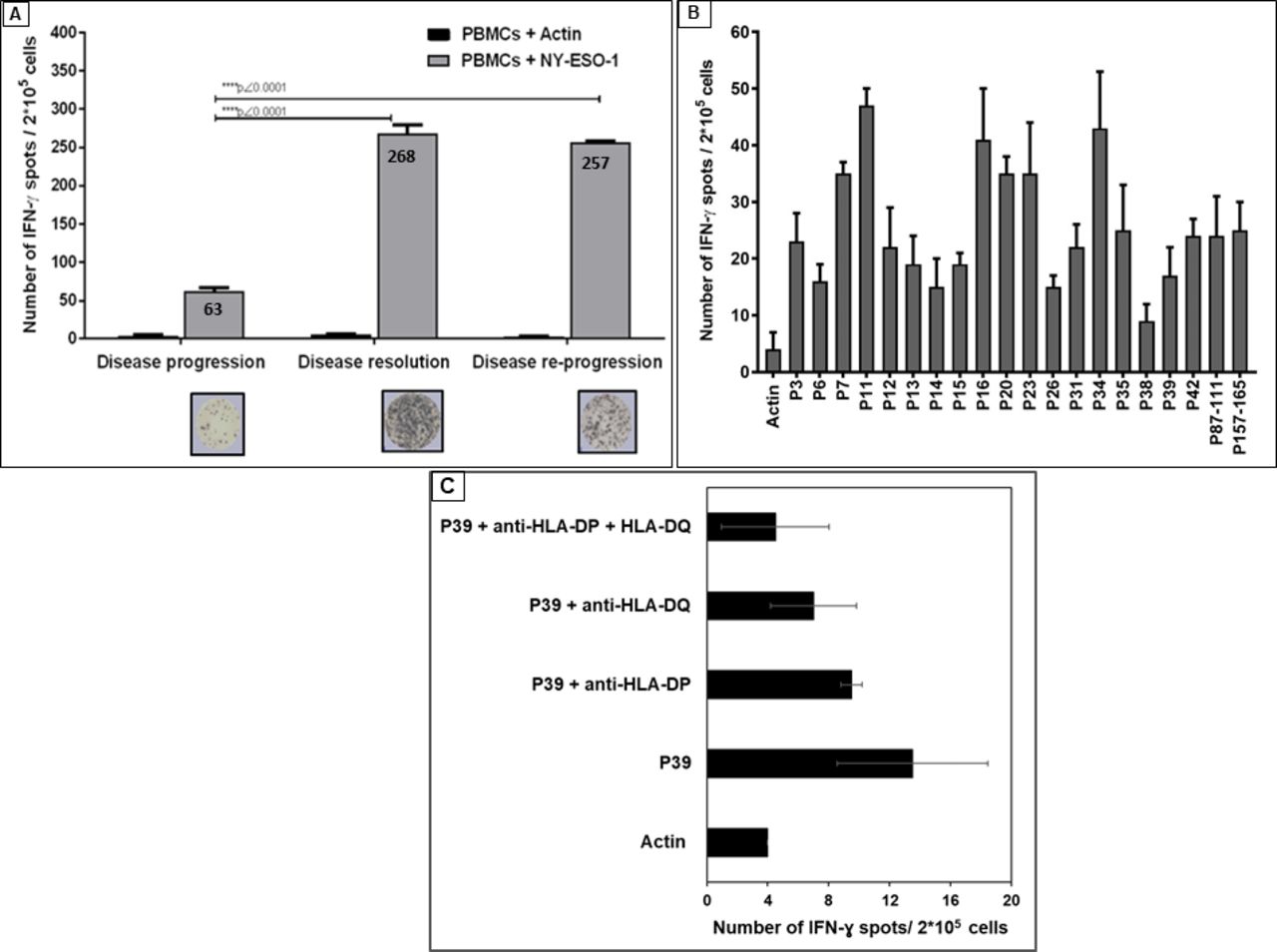
Enzyme-linked ImmunoSpot (ELISpot) results demonstrated that the NY-ESO-1-specific interferon-γ (IFN-γ) secretion by the patient T cells was detected at disease progression (after six cycles of first pembrolizumab treatment) against the NY-ESO-1 PepMix containing the whole 43 overlapping peptides (figure 2A). This IFN-γ secretion was significantly increased at both disease resolution (after concurrent RT/pembrolizumab treatment) and the disease re-progression using this NY-ESO-1 PepMix (****p˂0.0001) (figure 2A). Interestingly, 18 out of the 43 NY-ESO-1 single peptides tested (P3, P6, P7, P11, P12, P13, P14, P15, P16, P20, P23, P26, P31, P34, P35, P38, P39 and P42) were able to induce such specific IFN-γ production (online supplementary figure S2). This result was confirmed in a second experiment (figure 2B). Furthermore, a very high IFN-γ production (increased by sixfolds when compared with actin PepMix as a negative control) after stimulation with a promiscuous NY-ESO-1 peptide (P87–111), known to bind to multiple HLA-DR and HLA-DP4 alleles,16 was recorded (figure 2B). Similarly, a very high IFN-γ secretion (increased by sixfolds when compared with actin PepMix as a negative control) was also recorded when a known highly immunogenic NY-ESO-1/HLA-A-02:01 restricted epitope (P157–165)17 18 was used (figure 2B).

Enzyme-linked ImmunoSpot (ELISpot) assay for IFN-γ production by T cells from patient’s peripheral blood mononuclear cells (PBMCs). (A) IFN-γ production by patient’s T cells from PBMCs at disease progression, disease resolution and disease re-progression after in vitro challenge with NY-ESO-1 antigen (PepMix pool of 43 overlapping peptides for NY-ESO-1 antigen). (B) IFN-γ production by patient’s T cells from PBMCs at disease re-progression, in the presence of the selected 18 immunogenic single peptides (based on data presented in online supplementary figure S2) and two known immunogenic NY-ESO-1 peptides; the NY-ESO-1 promiscuous immunogenic 87–111 peptide (P87–111) and NY-ESO-1 HLA-A2-02:01 NY-ESO-1 restricted peptide (P157–165). For figure 2A, we performed unpaired non-parametric analysis of variance followed by Tukey’s multiple comparison test to compare T cell response at different stages of the patient’s disease (progression, resolution and re-progression). For figure 2B, we used non-parametric Mann-Whitney statistical analysis to compare the response to each peptide with the response to the negative control (actin). ELISpot assays were repeated twice with triplicates and the shown data corresponds to one representative experiment. P values ˂0.05 were considered statistically significant. (C). EliSpot assay showing the restriction response to HLA-DP and HLA-DQ molecules. IFN-γ, interferon-γ; NY-ESO-1, New York esophageal squamous cell carcinoma-1.
Identification of a novel promiscuous anti-NY-ESO-1 immunogenic CD4+ T cell epitope
Molecular tissue typing showed that the patient phenotype was HLA-A2-02:01 (online supplementary table 1). We used the Immune Epitope Database (IEDB1) prediction database to assess the patient’s HLA class II (DRB1*01:01, DRB1*13:03, DQA1*05:05, DQA1*01:01, DPA1*03:01, DPA1*01:03) binding predictability of the NY-ESO-1-derived peptides to these alleles (see Materials and Methods in the online supplemental file 1). Using this IEDB1 database, we have re-examined the binding affinity of the identified 18 NY-ESO-1 peptides that induced specific IFN-γ production response (figure 2B and online supplementary figure S2). We have demonstrated that 5 NY-ESO-1 immunogenic peptides (P23 (P89–103), P34 (P133–147), P38 (P149–163), P39 (P153–167), P42 (P165–179)) have a strong binding affinity to the different patient’s HLA class II alleles (online supplementary table 2). Interestingly, peptide P39 (P153–167) showed high binding affinity to both HLA-DQ and HLA-DP alleles (DQA1*05:05, DQA1*01:01, DPA1*03:01, DPA1*01:03). Moreover, this peptide (P39: LQQLSLLMWITQCFL) displayed a high binding affinity (data not shown) to the patient HLA-A2-02:01 allele and contains the known NY-ESO-1 HLA-A2-02:01 (P157–165) immunogenic epitope (SLLMWITQC)18 that has been also demonstrated in our report to induce specific IFN-γ secretion (figure 2B). To confirm the HLA-DP and DQ restriction element of P39, we used an antibody blocking experiment against these molecules. Anti-HLA-DP was able to inhibit 29.6% of IFN-γ production whereas anti-HLA-DQ resulted in 48.1% inhibition. Moreover, the combination of both antibodies was able to reduce IFN-γ production by 66.7% (figure 2C). Therefore, we have considered this NY-ESO-1 peptide (P39: P153–167) as a novel promiscuous immunogenic epitope restricted to different HLA alleles. Similarly, we used this IEDB1 database to re-examine the binding affinity of 11 subpeptides (each consists of 15 amino acids) derived from the known NY-ESO-1 promiscuous immunogenic peptide (P87–111)16 that consists of 25 amino acids (online supplementary table 3). Two predicted subpeptides, PP1 (P87–101) and PP2 (P88–102), showed high binding affinity to five different alleles of the patients (HLA-DR 01.01, HLA-DQ 01.01, HLA-DQ 05.05, HLA-DP 01.03 and HLA-DP 03.01) (online supplementary table 3). PP3 (P89–103) predicted to bind to the HLA-DR 01.01, HLA-DQ 05.05 and HLA-DP 01.03 alleles, whereas PP4 (P90–104) has the ability to bind to the HLA-DR 01.01 and HLA-DQ 05.05 alleles. Interestingly, 9 out of 11 subpeptides (PP1–PP9) also predicted to highly bind to the HLA-DQ 05.05 allele (online supplementary table 3).
Phenotyping and functional characterization of patient’s T cells
We first examined the CD4+ and CD8+ T cell distribution at different disease stages of the patient. Similar distribution of CD4+ and CD8+ T cells was recorded at disease progression and re-progression in which the CD4+ T cells were the most dominant population (55.1% CD4+ vs 36.6% CD8+ at disease progression and 56.4% CD4+ vs 34.4% CD8+ at disease re-progression) (figure 3A). However, the CD8+ T cell population was dominant (6.8% CD4+ vs 81% CD8+) at disease resolution (figure 3A). Because the patient expresses the HLA-A2-02:01 allele, we examined specific CD8+ T cells to the NY-ESO-1 HLA-A2-02:01 (P157–165) immunogenic epitope described above using an HLA-A2-NY-ESO-1-dextramer staining assay (Materials and Methods in the online supplemental file 1). CD8+/HLA-A2-NY-ESO-1-dextramer+ T cells were present at both disease progression (6.5%) and disease resolution (6.4%) and they were relatively increased at disease re-progression (14.5%) (figure 3B). CD8+ T cells from an HLA-A2+ and an HLA-A2− normal healthy controls stained negative for this dextramer (figure 3B). We next examined the cytotoxic T cells subset within the CD8+/HLA-A2-NY-ESO-1-dextramer+ T cells using the CD107 sensitive marker for the cytotoxic activity determination19 (Materials and Methods in the online supplemental file 1). Importantly, the CD107+ cytotoxic T cell subset was low at disease progression (2.5%), markedly increased at disease resolution (12.9%) and significantly decreased again at disease re-progression (3.6%) (figure 3C). CD8+ T cells from an HLA-A2+ and an HLA-A2− normal healthy controls stained negative for CD107 molecule (figure 3C). It has been reported that ICI blockade can enhance regulatory T cells (Tregs) inhibition.20 Similarly, it has been demonstrated that irradiated Tregs can exhibit a reduced capacity to suppress the proliferation of CD8+ T cells particularly when combined radioimmunotherapies are used.21 To this end, we analyzed the presence of the Tregs (CD4++ CD25high FOXP3+) population at disease resolution and disease re-progression in the peripheral blood mononuclear cells of the patient. Tregs distribution was low and comparable at both disease stages (online supplementary figure S3).
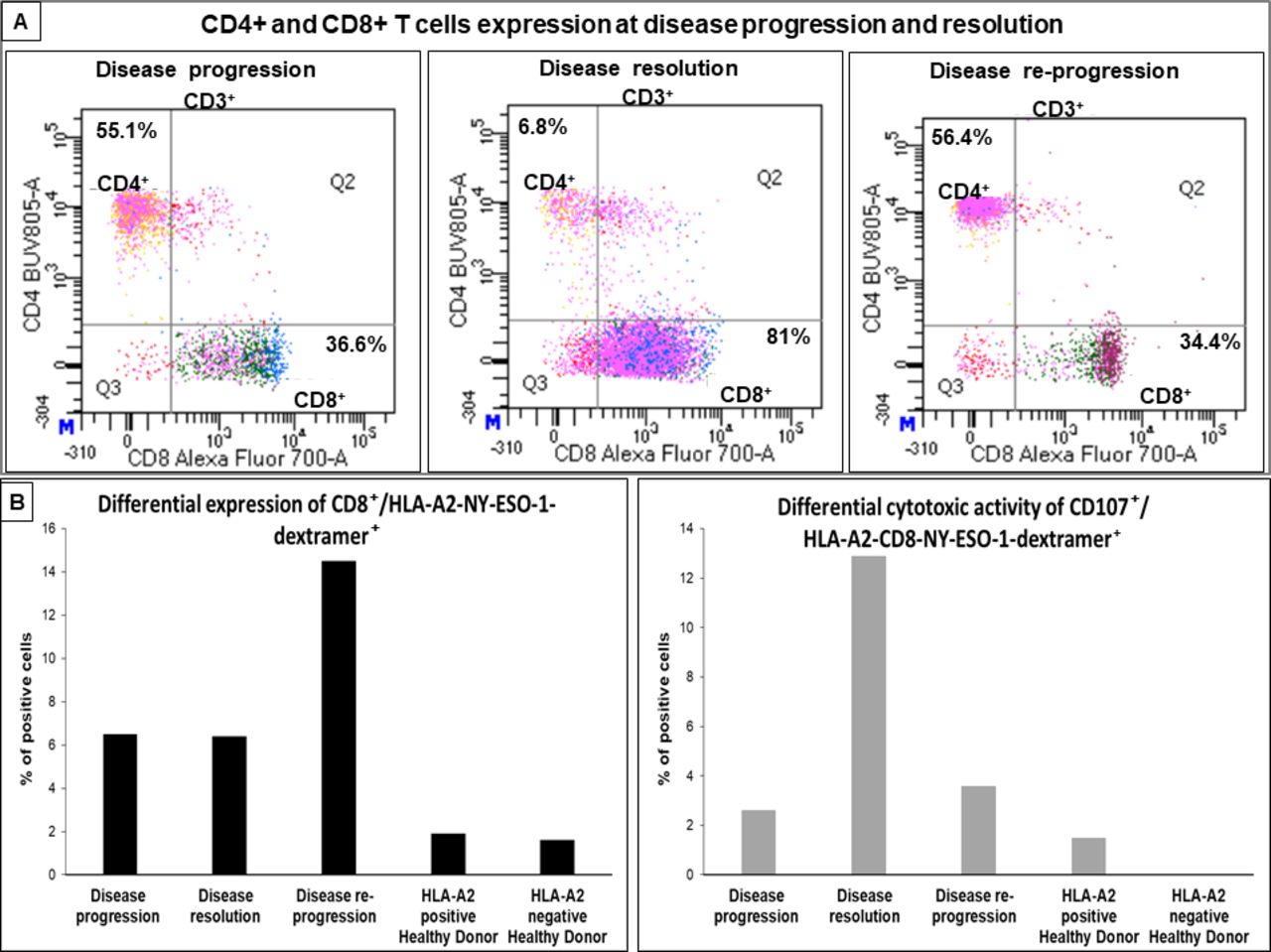
Phenotyping and functional characterization of patient’s T cells from peripheral blood using flow cytometry. (A) Dot blots represent CD4+ and CD8+ subsets (gated on CD3+ T cells) at disease progression, resolution and re-progression. CD4+ was the major T cell subset at disease progression and re-progression, but it dramatically decreased at resolution where we observed a significantly dominating CD8+ T cell population. (B) Histograms represent percentage of CD8+/HLA-A2-NY-ESO-1-dextramer+ T cell population in peripheral blood from the patient at disease progression, resolution and re-progression and from two healthy donors (HLA-A2 positive and negative donors). CD8+/HLA-A2-NY-ESO-1-dextramer+ T cells were relatively increased at disease re-progression. (C) Histograms represent percentage of CD107+ cytotoxic marker in CD8+/HLA-A2-NY-ESO-1-dextramer+ population in peripheral blood from the patient and the two healthy donors (HLA-A2 positive and negative donors). CD107+ cytotoxic CD8+/HLA-A2-NY-ESO-1-dextramer+ population was low at disease progression and re-progression and markedly increased at disease resolution. Flow cytometry assays were repeated two times and the shown data correspond to one representative experiment. NY-ESO-1, New York esophageal squamous cell carcinoma-1.
Differential expression of cytokines/chemokines in patient’s plasma
The cytokine/chemokine profile was investigated in the plasma of the patient at disease progression, disease resolution and disease re-progression using multiplex analysis (Materials and Methods in the online supplemental file 1). We classified the cytokines and/or chemokines profiles based on their upregulation or downregulation status during the three patient’s disease stages as mentioned above into two groups. Group 1 contains five cytokines (IFN-γ, tumor necrosis factor-α (TNF-α), IL-2, IL-5 and IL-6) that were present at disease progression, significantly downregulated at disease resolution and significantly upregulated again at disease re-progression (figure 4A). Group 2 contains four biomarkers (perforin, soluble FAS (sFAS), macrophage inflammatory protein-3α (MIP-3α) and C-X-C motif chemokine 11 (CXCL-11)/ITAC) that were present at disease progression, significantly upregulated at disease resolution and significantly downregulated again at disease re-progression (figure 4B).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Differential expression of cytokines/chemokines in patient’s plasma. The expression of different cytokines/chemokines was quantified by multiplexing assay in patient’s plasma from three time-points: first disease progression, disease resolution and re-progression. (A) Cytokines/chemokines decreased at disease resolution: the five biomarkers IFN-γ, TNF-α, IL-2, IL-5 and IL-6 were significantly downregulated after disease resolution and then upregulated once re-progression was observed. (B) Cytokines/chemokines increased at disease resolution: perforin, FAS, MIP-3α and CXCL-11 (ITAC) were significantly upregulated after disease resolution and then downregulated at re-progression. The experiments were repeated two times. In each experiment, the samples were done in triplicates. Values represent concentrations (mean±SD). Statistical analysis was carried out using non-parametric unpaired analysis of variance followed by Tukey’s multiple comparison test and p values ˂0.05 were considered statistically significant. CXCL-11, C-X-C motif chemokine 11; IFN-γ, interferon-gamma; IL-2, interleukin-2; IL-5, interleukin-5; IL-6, interleukin-6; MIP-3α, macrophage inflammatory protein-3α; sFAS, soluble FAS cytokine; TNF, tumor necrosis factor-alpha.
The remaining biomarkers (MIP-1α, MIP-1β, sFASL, sCD137, GM-CSF, IL-1 β, IL-4, IL-7, IL-8, IL-10, IL-12, IL-13, IL-17A, IL-21 and IL-23) were found to be upregulated only at disease progression (data not shown). Granzyme A and B were below detectable range (data not shown).
Discussion
We report here a patient with an advanced gastric MMR-deficient adenocarcinoma who was initially refractory to a PD-1 ICI but subsequently benefited from a long-lasting tumor control following the addition of a short course of localized RT. This disease evolution is suggestive of a primary resistance to anti-PD-1 checkpoint blockade that was overcome by the adjunction of RT. Sasaki et al 22 similarly recently reported dramatic responses to palliative RT after prior anti-PD-1 treatment in patients with metastatic gastric cancer, in line with the precept that RT may enhance the therapeutic action of immune checkpoint blockade.23–25 Several reports have provided evidence for a beneficial use of concurrent ICI/fractionated dose of RT in which the highest response rate was achieved if RT is given before immunotherapy.24 This may be accounted for by observations suggesting that RT has the potential to overcome the immunosuppressive effect of the TME in cold tumors,26 possibly through type I IFN signaling in dendritic cells (DCs) promoting the cross-priming of CD8+ T cells.27 On the contrary, it has been reported that ICI blockade can enhance Tregs inhibition.20 Similarly, it has been demonstrated that irradiated Tregs can exhibit a reduced capacity to suppress the proliferation of CD8+ T cells particularly when combined radioimmunotherapies are used.21 In the present report, Tregs distribution was low and comparable at disease resolution and disease re-progression in the peripheral blood of the patient. In line with our result, it has been demonstrated recently that Tregs with suppressive activity were upregulated on tumor-infiltrating T cells from patients with gastric cancer compared with their expressions on corresponding peripheral blood.28 Furthermore, it has been reported that an increased number of peripheral blood CD8+ T cells can predict a better response to RT in patients with cancer and suggested that antitumor immunity may play an important role in tumor regression by RT.29 We have demonstrated that CD8+ T cells were strikingly increased (81%) in the peripheral blood of the patient at disease resolution.
The patient presented in this report has MSI-H tumors that contain a high mutational load,4 which should result in an increased probability of immunogenic neoantigen expression that can predict the response to the anti-PD-1 treatment as shown in other studies.5 No predicted neoantigens were available to carry out such immunogenicity investigation in our report. Instead, we have used the NY-ESO-1 antigen to predict/monitor the immune response of the patient at different stages. The NY-ESO-1 antigen has been reported to be expressed in 20% of patients with metastatic gastric cancer8 and has the capacity to induce both natural antibody and T cell responses.9 Of note, RT has been shown to promote immunological recognition of cancer cells by increasing the expression of the NY-ESO-1 antigen.30 Moreover, anti-NY-ESO-1 immunity was associated with an improved clinical benefit to ICI.10 11 In the present report, we have shown an NY-ESO-1-specific IFN-γ secretion by the patient T cells appearing after six cycles of pembrolizumab treatment, which was significantly increased after combined radioimmunotherapy (****p˂0.0001) (figure 2A). Interestingly, as much as 18 out of the 43 NY-ESO-1 single peptides tested were immunogenic and were able to induce such specific IFN-γ production (figure 2B and online supplementary figure S2). However, only 5 out of the 18 NY-ESO-1 immunogenic peptides were predicted to have a strong binding affinity to different patient’s HLA class II alleles. It has been shown that the ability of peptides to induce immune response does not strongly correlate with their affinity for the HLA molecules.31 In addition, a peptide with a high binding affinity to an HLA molecule is not necessarily a good stimulator for T cell response.32 In this respect, we have found that P11 (P41–55), which is the most immunogenic peptide, has the lowest prediction binding ability. Whereas P38 (P149–163), which is the weakest immunogenic peptide, has a higher prediction binding ability (online supplementary table 2). Interestingly, we have identified a novel promiscuous CD4+ T cell immunogenic epitope P39 (P153–167: LQQLSLLMWITQCFL) restricted to all of the patient’s HLA-DQ and HLA-DP alleles (DQA1*05:05, DQA1*01:01, DPA1*03:01, DPA1*01:03), which also contains the NY-ESO-1 HLA-A2-02:01 CD8+ T cell immunogenic epitope (P157–165: SLLMWITQC).17 18 Furthermore, we have also confirmed the HLA-DQ and HLA-DP binding restriction of the novel P39 in this report. However, in vitro generation of anti-P39-specific T cell responses in HLA-DQ and HLA-DP matched cancer patients and healthy controls is needed to further support our current findings. Of note, the patient T cells had also a very high IFN-γ production against a known promiscuous NY-ESO-1 peptide (P87–111: LLEFYLAMPFTPMEAELARRSLAQ)16 as well as the highly immunogenic NY-ESO-1/HLA-A2-02:01 restricted epitope (P157–165: SLLMWITQC) (figure 2B).
The promiscuous NY-ESO-1 peptide identified in this report, containing both HLA class I/CD8+ and class II/CD4+-restricted T cell epitopes, may have contributed, along with other peptides, to the generation of an effective immune response in the patient. Indeed, it has been recently demonstrated that successful tumor rejection with ICI therapy requires CD4+ T cells, which act to recruit and activate CD8+ T cells, and this therapy was most effective when combined with a vaccine that contains epitopes for both CD8+ and CD4+ T cells.33 Gastric cancer has been shown to express HLA class II antigens.34 Interestingly, it has been demonstrated, in advanced gastric cancers, that the 5-year survival rate of patients with HLA-DR antigen-positive cancers was significantly higher compared with patients with HLA-DR antigen-negative cancers.34 Moreover, the expression of HLA class II-restricted tumor antigens is essential for a successful tumor rejection, and this indicates that CD4+ T cell activation must also occur in the TME.33 Nevertheless, it has been shown that ICI-induced antitumor immunity requires the activity of tumor antigens specific for both CD8+ and CD4+ T cells even in tumors that do not express HLA class II.33 In this respect, the RT treatment used in our report may not only help in the direct elimination of tumors and prevention of their progression but it also promote the upregulation and secretion of the NY-ESO-1 antigen by the cancer cells9 30 that was efficiently cross-presented by HLA class II on DCs to T cells leading to the specific antitumor responses in this patient.
We have demonstrated that specific CD8+ T cells to the NY-ESO-1 HLA-A2-02:01 (P157–165) immunogenic epitope were present in the peripheral blood of the patient 1 year after receiving RT/ICI therapy. However, when the cytotoxic T cell subset (CD107+/CD8+/HLA-A2-NY-ESO-1-dextramer+ T cells) was examined, the CD107-sensitive marker for the cytotoxic activity determination19 was low at disease progression, then significantly increased at disease resolution and dramatically decreased again at disease re-progression. Therefore, the CD107 molecule may be considered as a prognostic marker in this patient as it may contribute to the disease progression/re-progression seen in our report.
It has been shown that NY-ESO-1 antibody is frequently observed in the serum of patients with tumors expressing NY-ESO-1 but has not been detected in healthy controls and patients with NY-ESO-1 negative tumors.35 However, in the present report, we have observed very low levels of NY-ESO-1 antibody with no significant difference between the different stages of the disease (disease progression, disease resolution and disease re-progression). We have recently demonstrated that anti-PD-1 treatment can result in downregulation of NY-ESO-1 antibody expression in the plasma of patients with cancer.11 It is important to mention that all our samples tested for the NY-ESO-1 antibody detection, in the present report, were collected after several cycles of anti-PD-1 treatment and this may explain in part the very low levels of NY-ESO-1 antibody recorded here. Nevertheless, it has been shown that the NY-ESO-1 antibody frequency is very low in patients with advanced gastric cancer involving lymph nodes metastasis (21.7% out of 126 patients)8 and that NY-ESO-1 sero-negative patients can still present NY-ESO-1-specific CD8 T cells36 and this supports our observation in the present work.
Although we were not able to carry out an NY-ESO-1 staining in the patient tumor, the presence of such specific T cell immunity is a strong indication of pre-existence of the expression of the NY-ESO-1 antigen in the patient’s tumor.37
In this report, we have identified two groups of cytokines/chemokines that were differentially expressed in the plasma of the patient at different stages. The first group includes IL-2, IL-5, IFN-γ, TNF-α and IL-6 that were present at disease progression but significantly downregulated at disease resolution and then dramatically upregulated again at disease re-progression. IL-2 and IL-5 are usually produced, respectively, by Th1 and Th2 CD4+ T cells. Interestingly, we have shown that the number of CD4+ T cells was dramatically decreased (about eightfolds) in the peripheral blood of the patient at disease resolution and this may explain in part the very low production of these cytokines at this stage. In addition, the demonstration of the high significant increase of IL-2 at disease re-progression, observed in our report, is in agreement with a previous study, which showed a strong association between high levels of serum IL-2 and advanced stage of gastric cancer.38 The dramatic increase of IFN-γ in the plasma of the patient compared with its very low levels at disease resolution could be indicative of its role in establishing a proinflammatory microenvironment leading to re-progression of the disease. The demonstration of significantly higher levels of circulating blood IFN-γ in the advanced stages of patients with gastric cancer39 supports our findings. TNF-α is produced by malignant cells, leukocytes and other cells in the TME and may contribute to the development of the tissue architecture necessary for tumor growth and metastasis.40 Forones et al 38 have shown that patients with advanced stage of gastric cancer had significantly elevated serum levels of TNF-α and this was correlated with tumor progression. Similarly, we have shown significant high levels of TNF-α at both progression and re-progression of the disease. IL-6 is a pleiotropic cytokine that plays an important role in cell proliferation, survival, differentiation, migration and invasion. It regulates tumor progression and tumor metastasis by modulating tumor angiogenesis and tumor lympho-angiogenesis.41 It has been demonstrated that IL-6 may act on cancer cells through hepatocyte growth factor to promote and accelerate invasion as well as lymph node and hepatic metastasis in gastric cancer.42 The significant elevation of IL-6 levels recorded at disease re-progression in our report correlates well with other data demonstrating a strong association between high levels of serum IL-6 and unfavorable outcome in patients with gastric cancer.42 43 Finally, the observation of the decrease of cytokines associated with a protective Th1 polarized immunity (IFN-g, TNF-a and IL-2) at disease resolution and their increase at disease re-progression could be explained by the migration of such T cell population to the tumor site at resolution and their higher distribution in the peripheral blood at disease re-progression.
The second group contains CXCL-11/ITAC, MIP-3α, perforin and sFAS that were present at low levels at disease progression, then significantly increased at disease resolution and dramatically downregulated again at disease re-progression. The CXCL-11/ITAC chemokine is known to regulate immune cell migration, differentiation and activation.44 It has been demonstrated recently that treatment of patients with non-small cell lung cancer with docetaxel, an immune modulating drug, was able to induce CD8+ T cell recruitment to the TME by enhancing the secretion of CXCL-11 and improving the antitumor efficacy.45 In addition, CXCL-11 was also involved in eliciting a systemic immune response in these patients.45 Indeed, increased expression of CXCL-11 was positively correlated with prolonged overall survival of the patients.45 Other study showed that CXCL-11 modulates the immune-suppressive TME to an immune-supportive one and elucidates a potent and sustained systemic antitumor CD8+ T cell response with enhanced therapeutic efficacy and a survival benefit.46 Interestingly, we have shown that the number of systemic CD8+ T cells was significantly increased (about 2.3-folds) at disease resolution and this may explain in part the very high production of CXCL-11 at this stage. It has been shown that anti-PD-1 treatment could not only enhance T cell-mediated tumor regression but also increase the expression of CXCL-11.47 Interestingly, an animal study has demonstrated a critical correlation between CXCL-11/CXCR3-induced T cells homing to tumor site and anti-PD-1 treatment effect and suggested that anti-PD-1 therapy is not effective without the CXCL-11/CXCR3 axis.48 In this respect, the RT/anti-PD-1 treatment used in our report may increase the expression of CXCL-11, at disease resolution, that helped in the generation and activation of tumor-specific CD8+ T cells to directly eliminate tumors and prevent their progression. Very recently, upregulation of CXCL-11 was identified as a protective prognostic factor that significantly associated with overall survival of ovarian cancer.49 Indeed, it has been shown in other study that CXCL-11 occurred in the serum of patients with cancer 2 weeks post-treatment of anti-PD1/PDL-1 and sustained up to 6 weeks post-treatment.50 Interestingly, patients who developed immune-related adverse events had lower levels of CXCL-11 at baseline.50
MIP-3α, also known as CCL20, LARC or exodus, interacts with its specific CC-chemokine receptor 6 (CCR6) receptor, that is selectively expressed on DCs and memory T cells, and plays a major role in chemokine-mediated lymphocyte trafficking during gastric inflammation.51 52 It has been demonstrated that human TH17 cells express high levels of the CCR6 homing molecule that may be associated with their migration and retention within the tumors.53 Indeed, MIP-3α facilitates the trafficking of TH17 cells into tumors, which then mediate potent antitumor immunity by recruiting CD8+ T cells54 and DCs53 into the TME. We have shown in our report a significant increase in the expression of MIP-3α at disease resolution and this was dramatically decreased at disease re-progression. We have also demonstrated in our present work a significant increase in the expression of perforin at disease resolution and this was dramatically decreased at disease re-progression. Perforin-mediated Ag-specific tumor killing has been attributed to cytotoxic granule exocytosis from activated CD8+ T cells. It has been shown that newly produced perforin traffics directly to the immunological synapse, where it promotes cytotoxic activity.55 Interestingly, there was a correlation between rapid upregulation of perforin, produced by antigen-specific CD8+ T cells, and the expression of the cytotoxic CD107 marker.55 We have also shown in the present work a strong correlation between the cytotoxic T cells subset (CD107+/CD8+/HLA-A2-NY-ESO-1-dextramer+ T cells, figure 3C) and perforin production (figure 4B) at different stages of the disease (low at disease progression, very high at disease resolution and very low at disease re-progression). Finally, we have shown that sFAS was present at low levels at disease progression, then significantly increased at disease resolution and dramatically downregulated again at disease re-progression. It has been demonstrated recently that tumor resistance to ICI was linked to the ability of tumors to induce TIL apoptosis,56 which can be prevented by disturbing the FAS/FAS-ligand pathway.56 Interestingly, administration of sFAS was able to prevent TIL apoptosis and to increase the antitumor efficacy of adoptive T cell therapy and the efficacy of ICI.56 Furthermore, it has also been demonstrated that IFN-γ induces the inhibitory molecule FAS-L that contributed to the negative feedback of the immune response, and the use of IFN-γ-neutralizing antibody resulted in a strong reduction of apoptotic TIL.56 The dramatic increase of IFN-γ in the plasma of the patient at disease re-progression demonstrated in our report (figure 4A) may have contributed to the induction of FAS-L that resulted in TIL apoptosis, which may lead to the progression of the disease.
Conclusion
We have characterized the T cell immune response and analyzed the expression of plasma biological markers before and after radioimmunotherapy in a patient with a well-differentiated unresectable chemotherapy-resistant gastric adenocarcinoma. This patient showed complete tumor resolution 2 months after the term of the combined therapy with a subsequent long-lasting disease control before relapsing 12 months later with tumor re-progression and metastatic spread. Our study showed that radioimmunotherapy was able to enhance the immune response, which was found to correlate with disease evolution. The analysis of immunological markers in this patient showed a differential expression at different stages of the disease. Although we recognize one drawback of including only one patient in this current report, we suggest that our data would help in providing a critical understanding of important dynamic biomarkers of immune response useful for the prognosis of patients with cancer receiving combined radioimmunotherapy. Further studies are needed to be performed with larger number of patients with gastric adenocarcinoma treated with combined radioimmunotherapy to validate our findings.
Acknowledgments
We acknowledge Dr Mohamed Elkhalifa and Ms Zahra Abdi Ashur, from the histocompatibility and immunogenetics laboratory at Hamad Medical Corporation (HMC), for performing HLA typing on patient’s blood sample. We acknowledge Dr Prem Chandra for his help in the statistical analysis. We are grateful to the patient for his participation to the study and to the MRC at HMC for funding this project. We acknowledge Qatar National Library for supporting the publication.
References
Footnotes
Contributors SD designed and supervised the study. MMerhi, SD, ARaza, VPI and ARelecom drafted the manuscript. MMerhi, ARaza, VPI, KSS, DK and FS performed experiments and generated data. SM, SH and LAZ contributed to research experiments. NA and MJ coordinated the collection of patient samples. MMostafa, MAH, ARZG and MSEH provided clinical data. SU and MAH provided intellectual input and critical review of the manuscript. All authors contributed to manuscript revision, read and approved the submitted version.
Funding This case report was funded by Medical Research Center, Hamad Medical Corporation as part of MRC grant 17118/17.
Competing interests None declared.
Patient consent for publication Obtained.
Ethics approval The study was approved by the Institutional Review board committee of Hamad Medical Corporation, Doha, Qatar.
Provenance and peer review Not commissioned; externally peer reviewed.