Article Text

Abstract
Background Reinvigoration of T-cell exhaustion with antibodies has shown promising efficacy in patients with non-small-cell lung cancer (NSCLC). However, the characteristics of T-cell exhaustion with regard to tumor-infiltrating lymphocytes (TILs) are poorly elucidated in NSCLC. Here, we investigated the exhaustion status of TILs in NSCLC patients at the intraindividual and interindividual levels.
Methods We obtained paired peripheral blood, normal adjacent tissues, peritumoral tissues, and tumor tissues from 96 NSCLC patients. Features of T-cell exhaustion were analyzed by flow cytometry. T cells were categorized according to their programmed cell death-1 (PD-1) expression (PD-1high, PD-1int, and PD-1neg cells). Patients were classified based on the presence or absence of discrete PD-1high CD8+ TILs. Production of effector cytokines by CD8+ TILs was measured after T-cell stimulation with or without antibodies against immune checkpoint receptors.
Results Progressive T-cell exhaustion with marked expression of exhaustion-related markers and diminished production of effector cytokines was observed in PD-1high CD8+ TILs compared with PD-1int and PD-1neg CD8+ TILs. Patients with distinct PD-1high CD8+ TILs (PD-1high expressers) exhibited characteristics associated with a favorable anti-PD-1 response compared with those without these lymphocytes (non-PD-1high expressers). Combined inhibition of dual immune checkpoint receptors further restored effector cytokine production by CD8+ TILs following T-cell stimulation. PD-1high CD8+ T lymphocyte populations in the peripheral blood and tumors were significantly correlated.
Conclusions T-cell exhaustion was differentially regulated among individual patients and was prominent in a subgroup of NSCLC patients who may benefit from PD-1 blockade or combined blockade of other immune checkpoint receptors.
- lymphocytes
- tumor-infiltrating
- lung neoplasms
- programmed cell death 1 receptor
- tumor microenvironment
Data availability statement
Data are available on reasonable request. Data are available from the corresponding author HRK on reasonable request.
This is an open access article distributed in accordance with the Creative Commons Attribution Non Commercial (CC BY-NC 4.0) license, which permits others to distribute, remix, adapt, build upon this work non-commercially, and license their derivative works on different terms, provided the original work is properly cited, appropriate credit is given, any changes made indicated, and the use is non-commercial. See http://creativecommons.org/licenses/by-nc/4.0/.
Statistics from Altmetric.com
Background
T-cell exhaustion is a distinct type of T-cell dysfunction that develops during cancers and chronic infections.1 Exhausted T cells are characterized by progressive loss of effector functions, high and sustained expression of immune checkpoint inhibitory receptors, poor memory recall, metabolic dysregulation, and diminished homeostatic self-renewal.2 Of note, T-cell exhaustion is accompanied by transcriptional and epigenetic reprogramming during chronic T-cell receptor stimulation without antigenic clearance.3–7 Nowadays, targeting immune checkpoint receptors, including programmed cell death-1 (PD-1) and cytotoxic T-lymphocyte-associated protein-4 (CTLA-4), has been developed for reinvigorating exhausted T cells.8 9 In non-small-cell lung cancer (NSCLC), anti-PD-1 antibodies such as pembrolizumab and nivolumab used as monotherapies or as combination therapy with chemotherapy or other immune checkpoint inhibitors have substantially improved survival outcomes.10–16 However, it is noteworthy that only a minor proportion of patients derive therapeutic benefits from immune checkpoint inhibitors.17 18 Thus, further improvements in immunotherapeutic strategies based on an in-depth understanding of T-cell exhaustion are required to improve patient responses and outcomes.
Currently, expression of programmed death ligand-1 (PD-L1),19 20 spatial and temporal distribution of tumor-infiltrating lymphocytes (TILs),21 tumor mutation burden,22 gene expression profiles,23 and human leucocyte antigen heterogeneity24 have been suggested as biomarkers to predict the response to PD-1 blockade.25 In NSCLC, two key oncogenic alterations, epidermal growth factor receptor (EGFR) mutations and anaplastic lymphoma kinase (ALK) rearrangements, have been proposed to predict low response rates to anti-PD-1 treatment.20 26 However, only a few studies have evaluated the exhaustion of TILs, a direct target population of immune checkpoint blockade in the tumor microenvironment (TME), in patients with NSCLC. Investigation of T-cell exhaustion in tumor tissues from patients can provide mechanistic insights into the prediction of treatment response and resistance, as well as a basis for enhancing the therapeutic efficacy of combinatorial strategies.
In this study, we characterized the exhaustion of TILs in NSCLC by focusing on the expression of immune checkpoint receptors using freshly collected surgically resected tumor tissues. Further, we performed a correlation analysis of T-cell exhaustion with clinicopathological variables such as smoking status, histological subtype, alterations of EGFR or ALK, PD-L1 expression, and survival outcomes. Thus, we conceived an ex vivo model system to display anti-PD-1 responses in vivo and suggest a strategy for enhancing the therapeutic benefits of PD-1 blockade. Finally, we propose a peripheral blood-derived biomarker to predict T-cell exhaustion status in the TME.
Materials and methods
Specimen collection and preparation
Tissue specimens were collected at Yonsei Cancer Center (Seoul, Republic of Korea) from 96 patients with NSCLC who agreed to participate in the study from August 2017 to June 2021. Normal adjacent tissues (at least 5 cm away from gross tumor tissues) and peritumoral tissues (within 1 cm of gross tumor tissues) were obtained if required. Fresh tissue was cut into 1 mm3 pieces and digested with 1 mg/mL collagenase type IV (Worthington Biochemical, Lakewood, New Jersey, USA) and 0.01 mg/mL DNase I (Sigma-Aldrich, St. Louis, Missouri, USA) at 37°C for 20 min. The dissociated tissues were filtered through a 40 µm cell strainer (Corning, Corning, New York, USA) and washed with Roswell Park Memorial Institute-1640 medium (Corning) containing 10% fetal bovine serum (Biowest, FRA) and centrifuged. Tissue-infiltrating lymphocytes were enriched using a Percoll gradient (Sigma-Aldrich) followed by the staining of single-cell suspensions with the indicated fluorescent-conjugated antibodies.
Flow cytometry
Cells were stained with antibodies against cell-surface molecules for 20 min at 4°C. The LIVE/DEAD Fixable Red Dead Cell Stain Kit (Thermo Fisher Scientific, Waltham, Massachusetts, USA) was used to identify live cells. For intracellular staining, cells were fixed and permeabilized with the Foxp3 fixation/permeabilization solution (ThermoFisher Scientific) for 20 min at 4°C, and stained with antibodies against intracellular molecules for 20 min at 4°C. The stained cells were analyzed using FACS CytoFLEX (Beckman Coulter, Brea, California, USA) and the data were analyzed using FlowJo software V.10 (Treestar, San Carlos, California, USA). The antibodies used are listed in online supplemental table 1 and the gating strategies for flow cytometry are shown in online supplemental figure 1. Expression of PD-1 on CD8+ T lymphocytes from peripheral blood was used to gate for PD-1neg, PD-1int, and PD-1high populations.
Supplemental material
Supplemental material
Intracellular cytokine secretion assay
For the intracellular cytokine analysis of TILs, cells from dissociated tumors in the density range of 1×105 to 1×106 cells were cultured in a 96-well U-bottom plate containing 10 µg/mL of pembrolizumab (Selleckchem, Houston, Texas, USA), 10 µg/mL of ipilimumab (Selleckchem), soluble 50 ng/mL of anti-CD3 (BD Biosciences, San Jose, California, USA), and 50 ng/mL of anti-CD28 (BD Biosciences) for 24 hours at 37°C, followed by the addition of brefeldin A (BD Biosciences) and monensin (BD Biosciences) per the manufacturer’s protocol. To measure interferon (IFN)-γ and tumor necrosis factor (TNF) secretion levels by CD8+ TILs according to PD-1 expression on anti-CD3 and anti-CD28 stimulation, the pre-incubation step without brefeldin A and monensin for 24 hours was omitted. After 12 hours, the cells were harvested and stained. Intracellular cytokines were stained after surface staining, fixation, and permeabilization with Foxp3 fixation/permeabilization solution (Thermo Fisher Scientific).
Three-dimensional imaging of live cells
Three samples each from PD-1high and non-PD-1high expressers were used for live cell imaging analysis using t-stochastic neighbor embedding (t-SNE), a tool for visualizing high-dimensional data. Each sample was down-sampled to 20,000 randomly selected CD3+ live and singlet-gated cells. A total of 60,000 cells were merged. Then, 53,574 cells among 120,000 total cells were gated for CD8+ and visualized using t-SNE for analysis using FlowJo software V.10.6.2 (Treestar) with 3,000 iterations and a perplexity parameter of 100.
PD-L1 expression in tumor tissues
PD-L1 expression in the archived specimens was determined by immunohistochemical analysis using anti-PD-L1 antibodies (clone SP263). The tumor proportion score was determined for the positivity of PD-L1 expression on tumor cells as described previously.27 PD-L1 expression in freshly collected surgically resected tumor tissues was determined by flow cytometry.
Statistical analysis
Statistical significance was assessed using two-tailed Student’s t-test, χ2 test, or one-way analysis of variance with Tukey’s correction, as appropriate, with the Prism .7.0 software (GraphPad). Values of p≤0.05 (*), ≤0.01 (**), ≤0.001 (***), and ≤0.0001 (****) were considered significant.
Results
CD8+ TILs from NSCLC patients were subdivided into three distinct subpopulations based on PD-1 expression
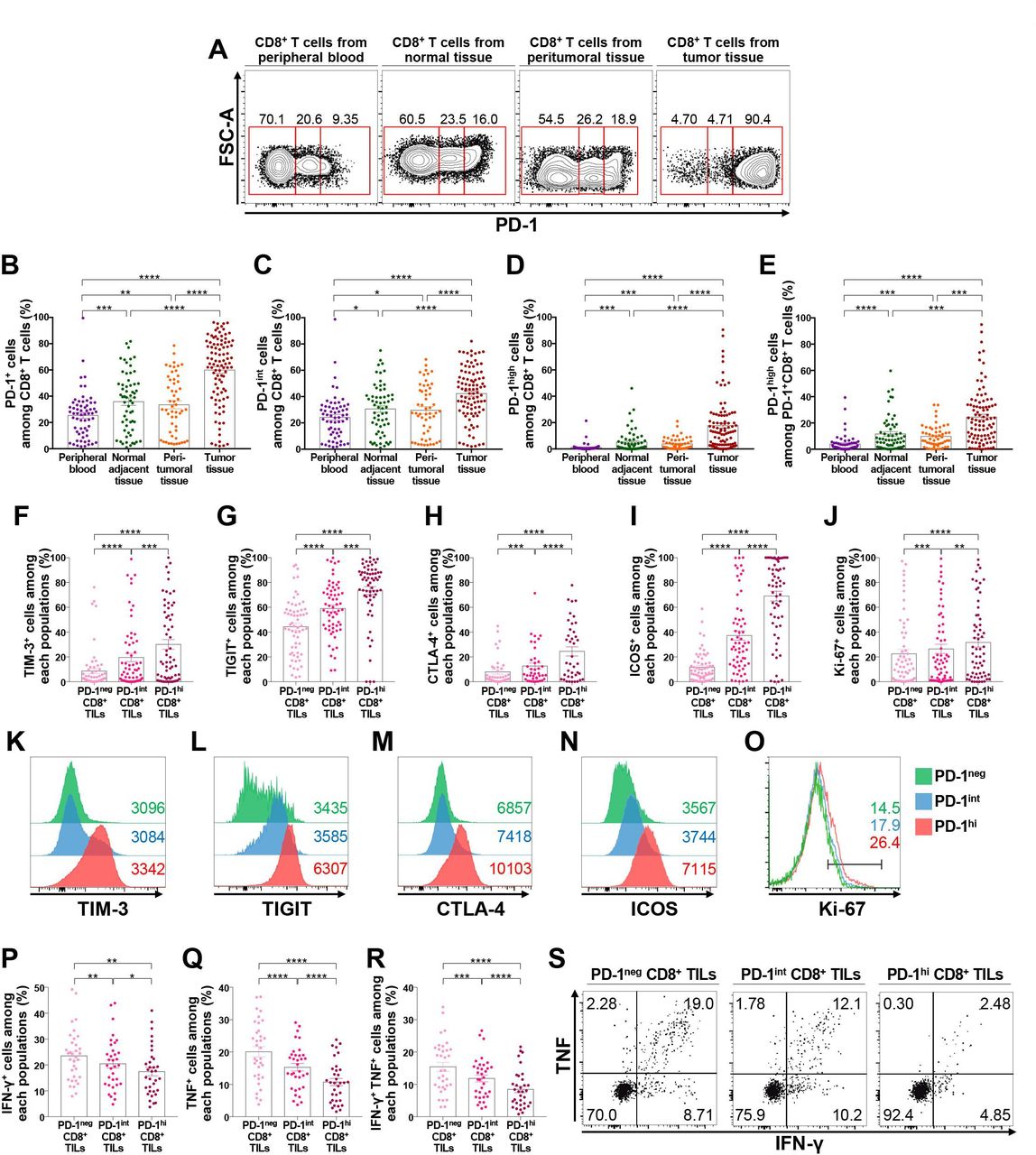
First, we examined PD-1 expression on CD8+ T lymphocytes from the peripheral blood, normal adjacent tissues, peritumoral tissues, and tumor tissues of patients with NSCLC (figure 1A). The frequency of PD-1+ cells among CD8+ T lymphocytes was the highest in CD8+ TILs, followed by CD8+ T lymphocytes from peritumoral tissues, normal adjacent tissues, and peripheral blood (figure 1B). We then subdivided CD8+ T lymphocytes into three distinct populations based on PD-1 expression: PD-1neg, PD-1int, and PD-1high (figure 1A). PD-1neg CD8+ T lymphocytes constituted the major population of CD8+ T lymphocytes from peripheral blood samples, whereas PD-1high CD8+ T lymphocytes were relatively enriched in CD8+ TILs (figure 1B–E). Notably, PD-1+ cells in CD8+ T lymphocytes from peripheral blood samples were mostly distributed in PD-1int populations (figure 1C). Importantly, the frequency of PD-1high CD8+ T cells was positively correlated among the normal adjacent tissues, peritumoral tissues, and tumor tissues (online supplemental figure 2), indirectly supporting field cancerization.28
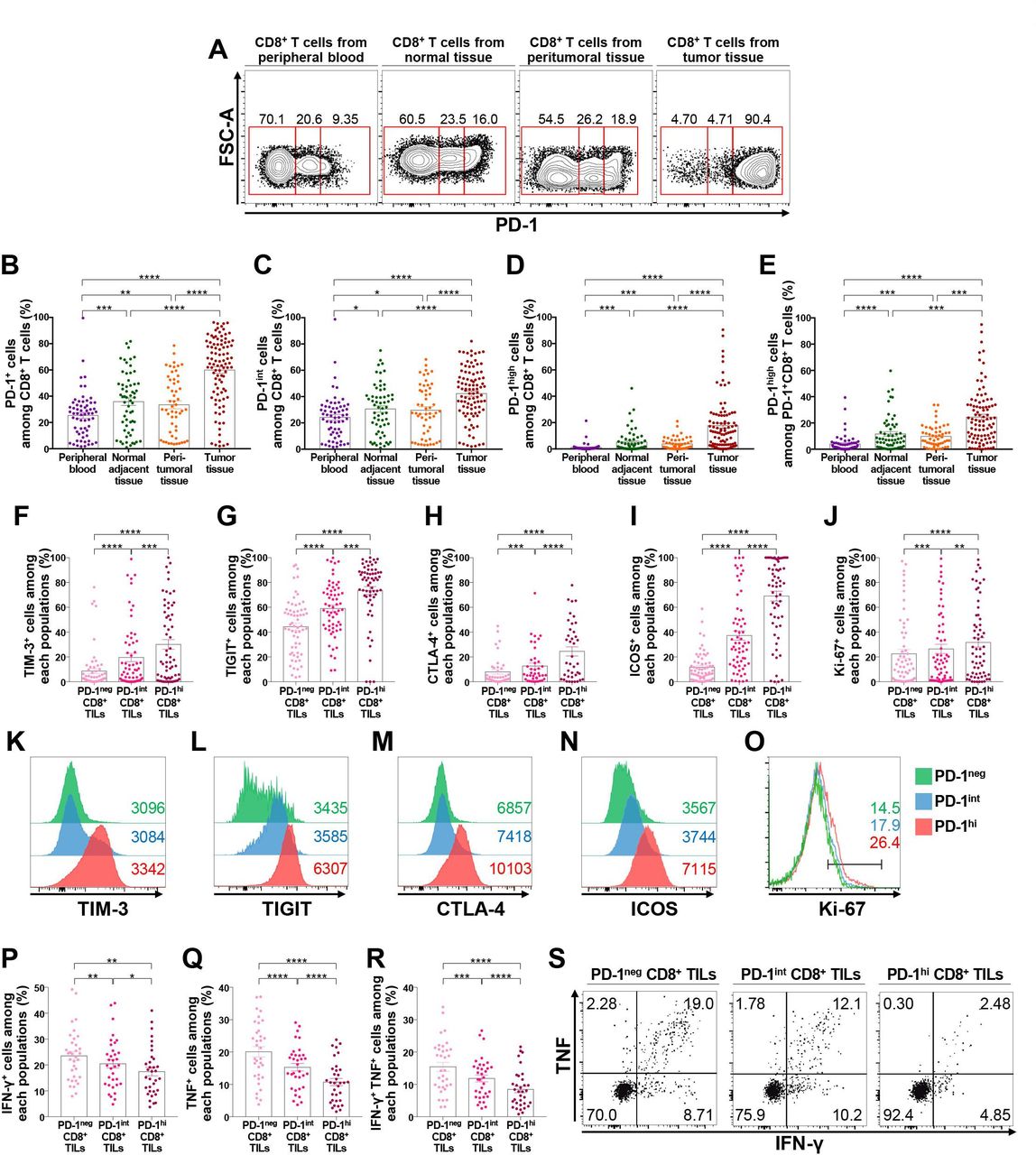
PD-1high CD8+ T lymphocytes are enriched in the tumor-immune microenvironment and highly express exhaustion-related and activation-related molecules. (A–E) Expression of PD-1 in CD8+ T lymphocytes from the peripheral blood, normal adjacent tissue, peritumoral tissue, and tumor tissue from NSCLC patients. Representative flow cytometry plots (A); frequencies of PD-1+ (B), PD-1int (C), and PD-1high (D) cells among CD8+ T lymphocytes; and frequency of PD-1high cells among PD-1+CD8+ T lymphocytes (E). (F–O) Expression of exhaustion-related and activation-related molecules on CD8+ TILs according to PD-1 expression. Representative histogram and the frequency of TIM-3+ (F, K), TIGIT+ (G, L), CTLA-4+ (H, M), ICOS+ (I, N), and Ki-67+ (J, O) cells among CD8+ TILs. (P–S) Production of IFN-γ (P), TNF (Q), and both IFN-γ and TNF (R) of CD8+ TILs on stimulation with anti-CD3 and anti-CD28, and a representative flow cytometry plot (S). *p<0.05; **p<0.01; ***p<0.001; ****p<0.0001. CTLA-4, cytotoxic T-lymphocyte-associated protein-4; FSC-A, forward scatter area; ICOS, inducible T-cell costimulator; IFN-γ, interferon-γ; NSCLC, non-small-cell lung cancer; PD-1, programmed cell death-1; TIGIT, T cell immunoreceptor with Ig and ITIM domains; TILs, tumor-infiltrating lymphocytes; TIM-3, T cell immunoglobulin and mucin domain-containing protein 3; TNF, tumor necrosis factor.
To further understand the characteristics of CD8+ TILs distinguished by differential PD-1 expression, we investigated the expression pattern of other immune checkpoint receptors such as T cell immunoglobulin and mucin domain-containing protein 3 (TIM-3), T cell immunoreceptor with Ig and ITIM domains (TIGIT), and cytotoxic T-lymphocyte-associated protein-4 (CTLA-4) in PD-1high, PD-1int, and PD-1neg CD8+ TILs (figure 1F–H and K–M). Notably, the expression of these immune checkpoint inhibitory receptors was predominantly observed in the PD-1high population, suggesting more progressed T-cell exhaustion features in this population. Expression of the costimulatory molecule inducible T-cell costimulator (ICOS) and the proliferation marker Ki-67 was also higher in the PD-1high population than in the PD-1int and/or PD-1neg populations (figure 1I,J and N,O), suggesting the indispensable relationship between T-cell activation and exhaustion.29 Of note, tissue-resident memory CD8+ T lymphocytes co-expressing CD39 and CD103 were significantly enriched in PD-1high populations (online supplemental figure 3), consistent with previous reports.30 31 The same results were observed when we examined tumor-infiltrating FoxP3– non-Treg and FoxP3+ Treg CD4+ T lymphocytes for the percentages of TIM-3+, TIGIT+, CTLA-4+, ICOS+, and Ki-67+ cells based on their PD-1 expression (online supplemental figures 4 and 5). In addition, there was a strong correlation between the frequency of PD-1high or PD-1+ cells among tumor-infiltrating CD8+, FoxP3– non-Treg, and FoxP3+ Treg CD4+ T lymphocytes (online supplemental figure 6), revealing the shared machinery of PD-1 induction between different T cell populations.32 Intracellular cytokine staining showed that the population producing effector cytokines was smaller in PD-1high CD8+ T cells than in PD-1int and PD-1neg cells on stimulation with anti-CD3 and anti-CD28 (figure 1P–S). Taken together, these results indicate that the T-cell exhaustion process is largely limited to the TME, and that PD-1high CD8+ TILs present features of more progressed T-cell exhaustion than PD-1int and PD-1neg T lymphocytes, as indicated by the prominent expression of exhaustion-related markers and diminished effector cytokine production, consistent with previous literature.33 34
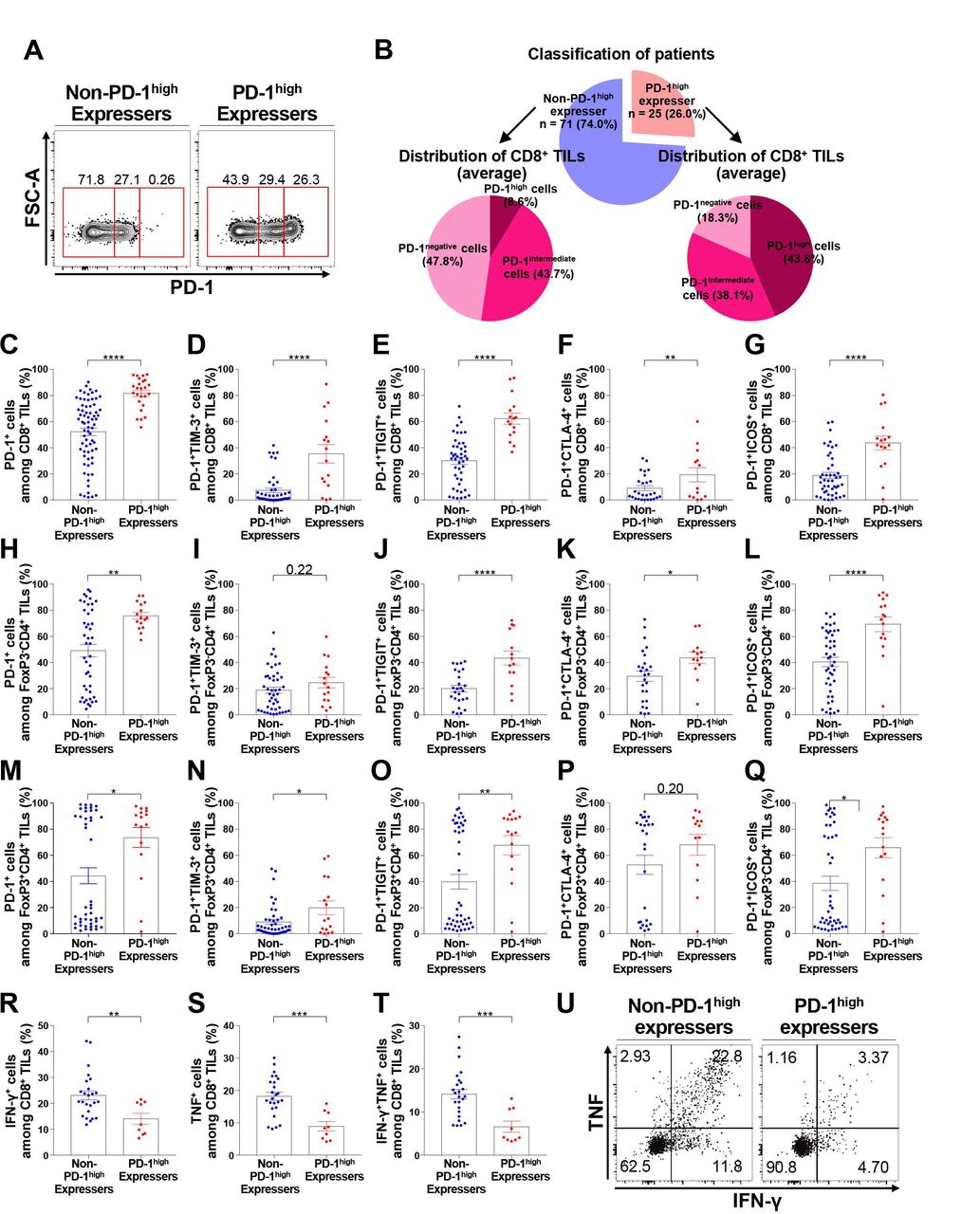
Patients with NSCLC can be classified into two subgroups based on the presence of distinct PD-1high CD8+ TILs
Based on the presence or absence of distinct PD-1high subpopulations in CD8+ TILs, we identified two distinct patient subgroups (figure 2A), namely PD-1high expressers (presence of distinct PD-1high CD8+ TILs) and non-PD-1high expressers (absence of distinct PD-1high CD8+ TILs), via flow cytometric analysis. Consequently, 25 PD-1high expressers (26.0%) and 71 non-PD-1high expressers (74.0%) comprised our cohort (figure 2B). The frequency of PD-1+ cells among CD8+ TILs was higher in PD-1high expressers than in non-PD-1high expressers (figure 2C). Furthermore, the frequency of PD-1+TIM-3+, PD-1+TIGIT+, PD-1+CTLA-4+, and PD-1+ICOS+ cells among CD8+ TILs was significantly higher in PD-1high expressers than in non-PD-1high expressers (figure 2D–G). Similar results were observed in tumor-infiltrating FoxP3– non-Treg and FoxP3+ Treg CD4+ T lymphocytes (figure 2H–Q), further confirming the close relationship between the exhaustion process of different T-cell subsets in the TME. In terms of effector cytokine production, the frequency of IFN-γ+ and/or TNF+ cells was lower in PD-1high expressers than in non- PD-1high expressers (figure 2R–U). Analysis with the t-SNE algorithm showed that the expression of immune checkpoint molecules, including PD-1, TIM-3, TIGIT, CTLA-4, and ICOS, was evidently higher in CD8+ TILs from PD-1high expressers than in those from non-PD-1high expressers (online supplemental figure 7). In line with this observation, the levels of various immune checkpoint inhibitory receptors were positively correlated with each other in bulk tumor tissue samples from The Cancer Genome Atlas NSCLC cohort (online supplemental figure 8), supporting the justification of the combined blockade of immune checkpoint inhibitory receptors in PD-1high expressers.

Distinct subgroups of NSCLC patients according to the presence of discrete PD-1high CD8+ TILs. (A) Representative flow cytometry plots showing the two subgroups of NSCLC patients, namely PD-1high expressers and non-PD-1high expressers. (B) Classification of patients and average distribution of PD-1 expression in CD8+ TILs in each subgroup. (C–G) Frequencies of PD-1+ (C), PD-1+TIM-3+ (D), PD-1+TIGIT+ (E), PD-1+CTLA-4+ (F), and PD-1+ICOS+ (G) cells among CD8+ TILs in non-PD-1high expressers and PD-1high expressers. (H–L) Frequencies of PD-1+ (H), PD-1+TIM-3+ (I), PD-1+TIGIT+ (J), PD-1+CTLA-4+ (K), and PD-1+ICOS+ (L) cells among tumor-infiltrating FoxP3– CD4+ T lymphocytes in non-PD-1high expressers. (M–Q) Frequencies of PD-1+ (M), PD-1+TIM-3+ (N), PD-1+TIGIT+ (O), PD-1+CTLA-4+ (P), and PD-1+ICOS+ (Q) cells among FoxP3+CD4+ TILs in non-PD-1high expressers and PD-1high expressers. (R–U) Production of IFN-γ (R), TNF (S), and both IFN-γ and TNF (T) of CD8+ TILs from non-PD-1high expressers and PD-1high expressers on stimulation with anti-CD3 and anti-CD28, and a representative flow cytometry plot (U). *p<0.05; **p<0.01; ***p<0.001; ****p<0.0001. CTLA-4, cytotoxic T-lymphocyte-associated protein-4; FoxP3, forkhead box protein P3; FSC-A, forward scatter area; ICOS, inducible T-cell costimulator; IFN-γ, interferon-γ; NSCLC, non-small-cell lung cancer; PD-1, programmed cell death-1; TIGIT, T cell immunoreceptor with Ig and ITIM domains; TILs, tumor-infiltrating lymphocytes; TIM-3, T cell immunoglobulin and mucin domain-containing protein 3; TNF, tumor necrosis factor.
Clinicopathological characteristics and survival outcomes of PD-1high expressers and non-PD-1high expressers
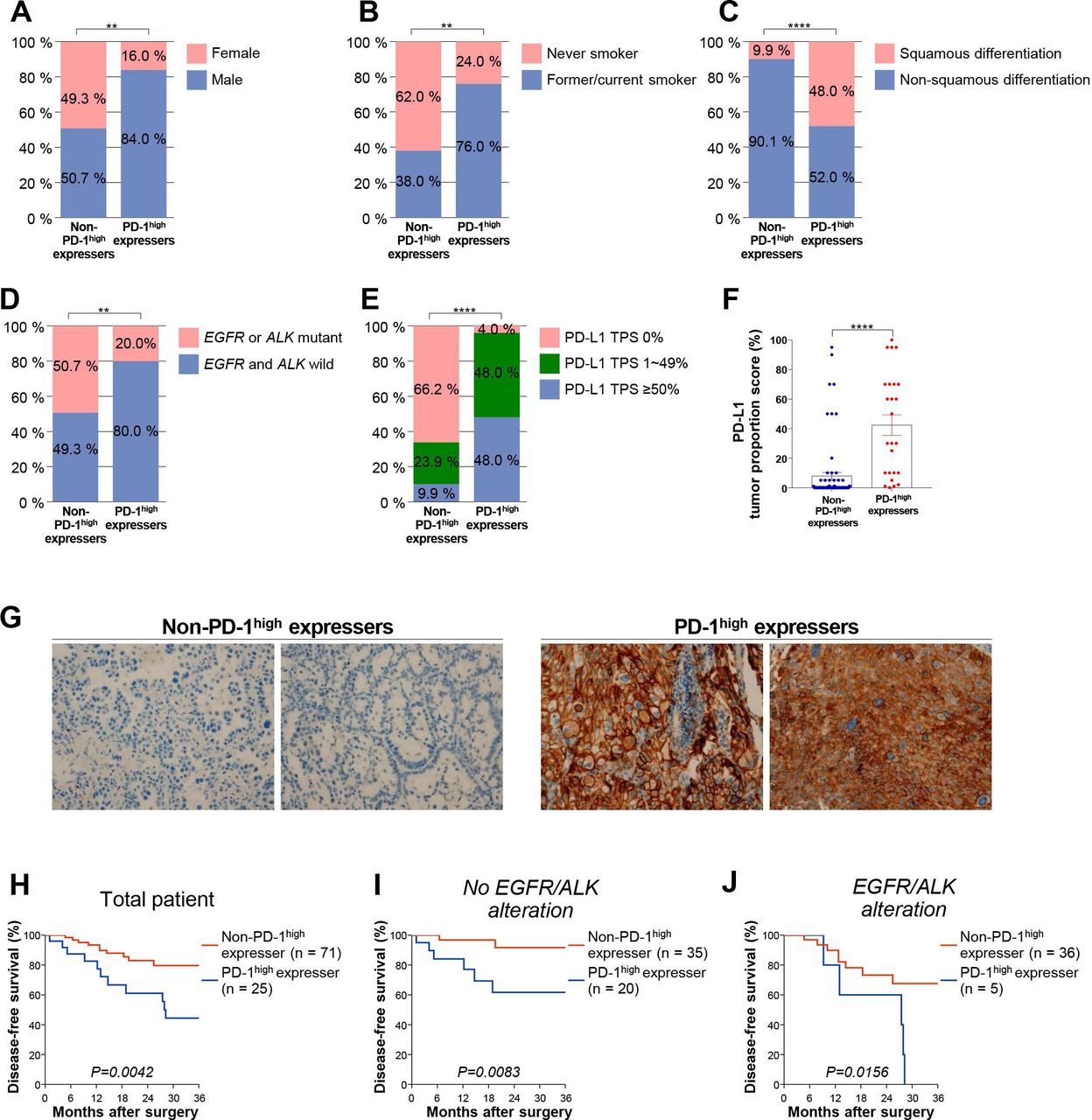
PD-1high expressers and non-PD-1high expressers did not significantly differ with regard to age and stage. However, PD-1high expressers were more likely to be males with a smoking history and a squamous histology, and had EGFR and ALK wild-type tumors (table 1, figure 3A–D). Based on the analysis revealing that EpCAM+CD45- cells were major PD-L1 carriers in NSCLC (online supplemental figure 9), we further explored PD-L1 expression on tumor cells via immunohistochemistry. PD-L1 expression on tumor cells was significantly higher in PD-1high expressers than in non-PD-1high expressers both in terms of positivity (cut-off of 1% and 50%, respectively) and tumor proportion score (figure 3E–G), although prediction of PD-1high expressers based on PD-L1 expression was incomplete (online supplemental figure 10). Moreover, PD-1high expressers had a poorer prognosis than non-PD-1high expressers with respect to disease-free survival after surgical resection (figure 3H). In addition, univariate and multivariate analyses after adjustment for alteration in EGFR or ALK and PD-L1 positivity confirmed that the presence of distinct PD-1high CD8+ TILs independently predicts poor survival outcomes (online supplemental table 2). When subgroup analysis was performed according to the presence or absence of alterations in EGFR or ALK, this significant association was maintained (figure 3I and J). Intriguingly, the effector cytokine-producing capacity was lower in patients with a smoking history and in tumors with squamous differentiation and/or without alterations in either EGFR of ALK (online supplemental figure 11). Furthermore, diminished functionality of CD8+ TILs was associated with worse disease-free survival (online supplemental figure 11). Taken together, these results indicate that classification based on the PD-1 expression of CD8+ TILs facilitates the identification of a patient subgroup associated with a worse prognosis. In addition, this subgroup of patients likely exhibits the clinicopathological characteristics (eg, high PD-L1 expression, EGFR and ALK wild-type, and previous history of smoking) known to be associated with the benefits from anti-PD-1-based immunotherapy.20 25 35 36

Clinicopathological variables and survival outcomes in non-PD-1high expressers and PD-1high expressers. (A) Distribution of sex, (B) smoking status, (C) histology, (D) presence of oncogenic EGFR or ALK mutation, and (E) PD-L1 positivity (≥1% and ≥50% as a tumor proportion score) in non-PD-1high expressers and PD-1high expressers. (F) PD-L1 expression as a tumor proportion score in non-PD-1high expressers and PD-1high expressers. (G) Representative immunohistochemistry images of PD-L1 from two non-PD-1high expressers and PD-1high expressers, respectively. (H–J) Disease-free survival of non-PD-1high expressers and PD-1high expressers for all patients (H; n=96), patients with wild-type EGFR and ALK (I; n=55), and patients with mutant-type EGFR or ALK (J; n=41). **p<0.01; ****p<0.0001. ALK, anaplastic lymphoma kinase; EGFR, epidermal growth factor receptor; PD-1, programmed cell death-1; PD-L1, programmed death ligand-1; TPS, Tumor Proportion Score.
Baseline characteristics of patients
Functionality of CD8+ TILs from NSCLC patients is further restored after combined immune checkpoint blockade with anti-PD-1 and anti-CTLA-4 antibodies
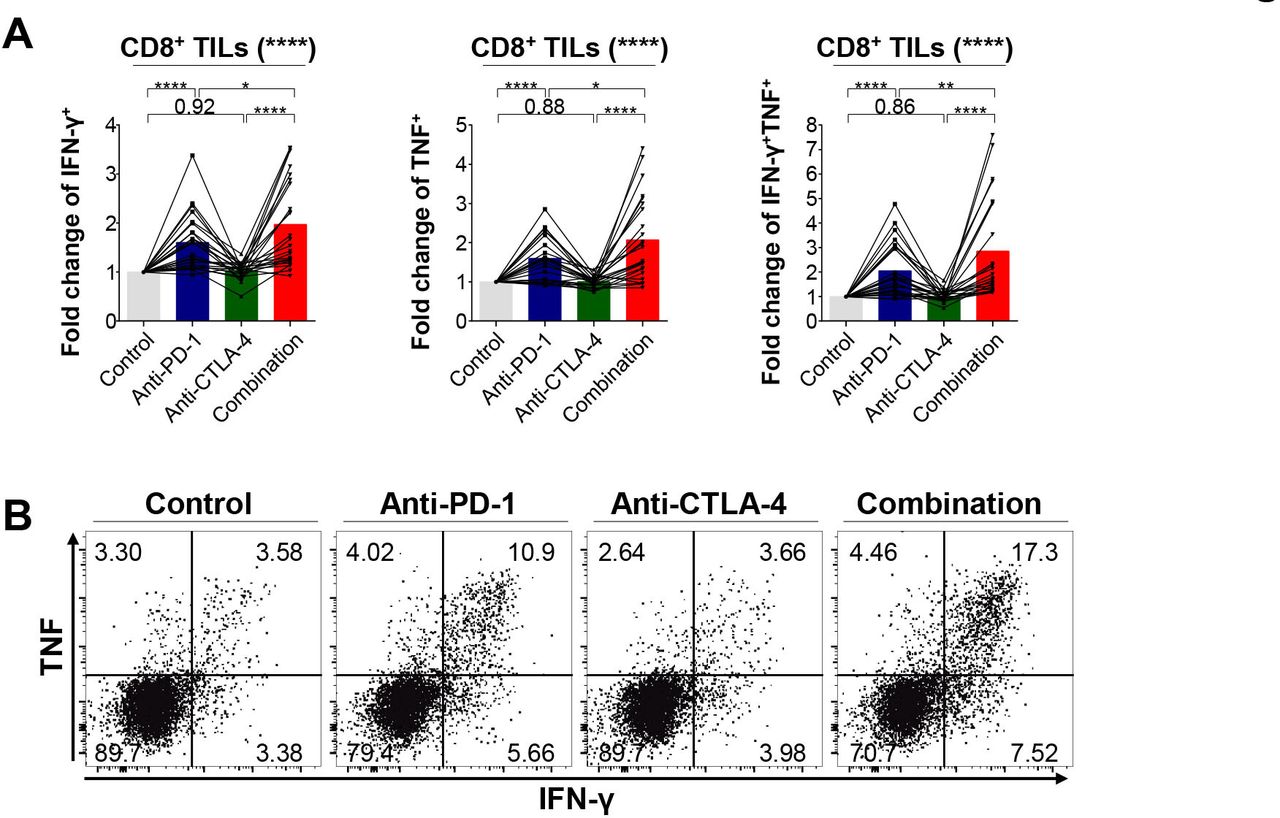
Since most CD8+ TILs from PD-1high expressers were more likely to express multiple immune checkpoints, we examined whether a PD-1 inhibitor in conjunction with a CTLA-4 inhibitor cooperatively restores the function of CD8+ TILs. First, we tested whether the functions of CD8+ TILs are enhanced by ex vivo single blockade of PD-1 or CTLA-4. Hence, we evaluated the production of IFN-γ and TNF by CD8+ TILs on anti-CD3 and anti-CD28 stimulation in the presence or absence of anti-PD-1 or anti-CTLA-4 antibodies (figure 4A,B). This functional analysis showed that PD-1 blockade alone increased the capacity of CD8+ TILs to produce IFN-γ, whereas CTLA-4 blockade alone was not as effective. More importantly, the combined PD-1 and CTLA-4 blockade significantly enhanced the polyfunctionality of CD8+ TILs to produce IFN-γ and TNF concurrently on anti-CD3/28 stimulation compared with PD-1 or CTLA-4 blockade alone, suggesting a potential synergism of this combination as witnessed in the CheckMate-227 study.13 This synergism was more evident in PD-1high expressers than in non-PD-1high expressers (online supplemental figure 12). Collectively, these results indicate that the combined blockade of immune checkpoint inhibitory receptors further restores the polyfunctionality of CD8+ TILs compared with monotherapy, consistent with previous studies.37 38
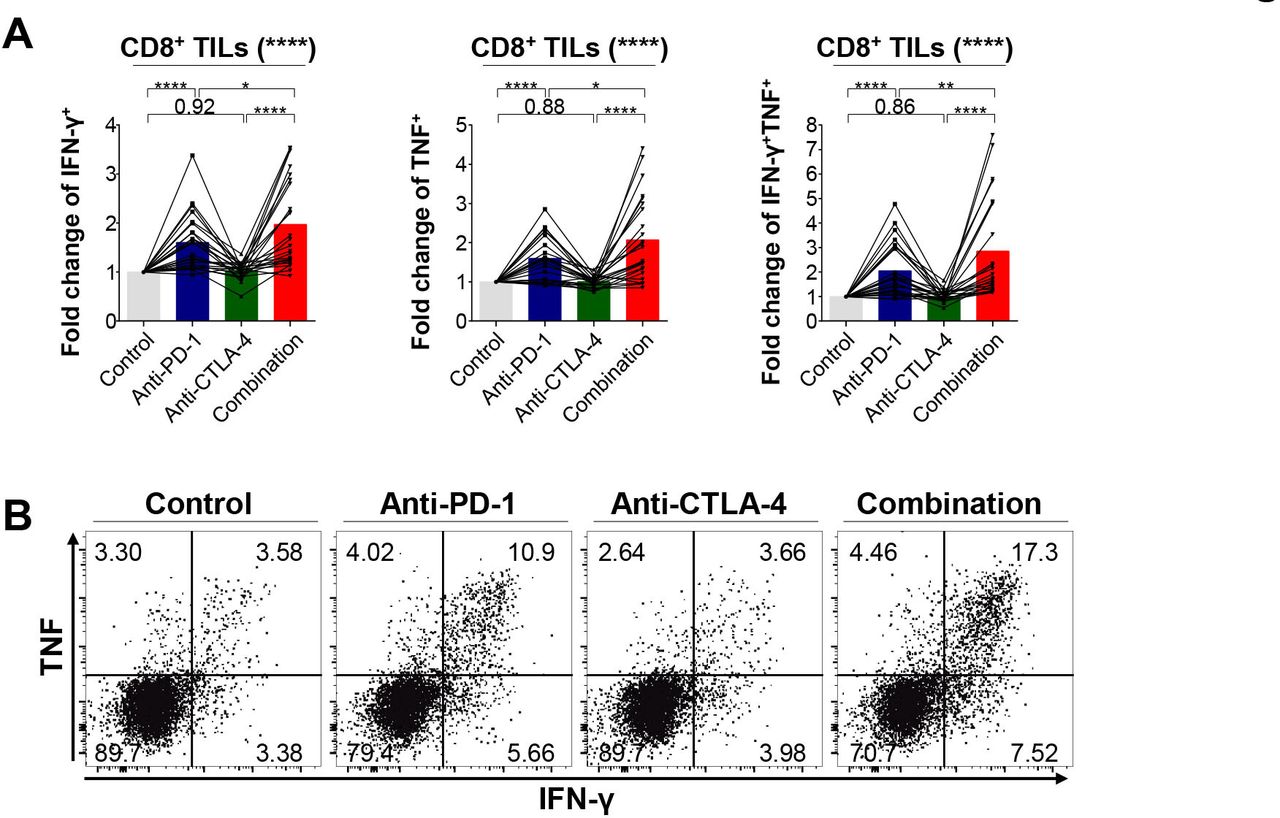
Efficacy of single PD-1 blockade and combined blockade of PD-1 and CTLA-4 on the effector cytokine production of CD8+ TILs (n=24). (A–B) Fold change of IFN-γ+, TNF+, and IFN-γ+TNF+ cells among CD8+ TILs relative to isotype controls on stimulation (A) and representative flow cytometry plot (B). *p<0.05; **p<0.01; ****p<0.0001. CTLA-4, cytotoxic T-lymphocyte-associated protein-4; IFN-γ, interferon-γ; PD-1, programmed cell death-1; TILs, tumor-infiltrating lymphocytes; TNF, tumor necrosis factor.
Frequency of PD-1high CD8+ T lymphocytes in peripheral blood predicts two subgroups of NSCLC patients
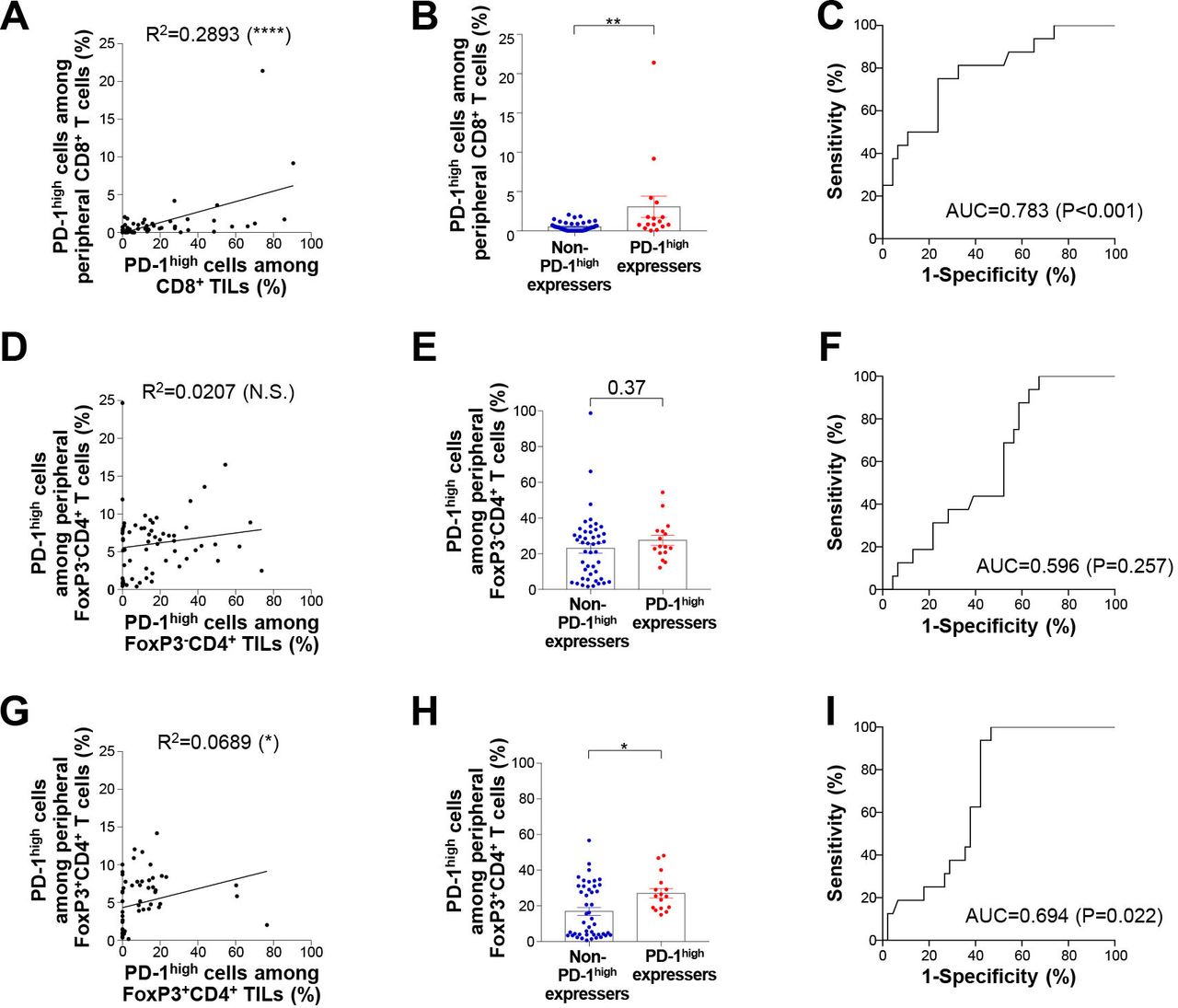
We next focused on PD-1high CD8+ T lymphocytes in the peripheral blood to examine whether the frequency of these cells can help identify subgroups of NSCLC patients. A significant correlation was found in the plot of the frequency of PD-1high CD8+ T lymphocytes from the peripheral blood and tumor (figure 5A). Notably, the frequency of PD-1+CD8+ T lymphocytes from the peripheral blood was significantly higher in PD-1high expressers than in non-PD-1high expressers (figure 5B). Moreover, the frequency of circulating PD-1high CD8+ T lymphocytes predicted PD-1high expressers and non-PD-1high expressers fairly well (figure 5C). Similar findings were obtained in the analysis of tumor-infiltrating FoxP3+ Treg CD4+ T lymphocytes but these effects were not recapitulated in the case of tumor-infiltrating FoxP3– non-Treg CD4+ lymphocytes (figure 5D–I). Overall, these results further support the potential usefulness of analyzing PD-1high populations from peripheral blood-derived T lymphocytes in the absence of tumor tissues.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Two subgroups of NSCLC patients can be distinguished by the frequency of peripheral PD-1+ T lymphocytes. (A) Correlation between PD-1high cells among CD8+ TILs and PD-1high cells among peripheral CD8+ T lymphocytes. (B) Frequency of PD-1high cells among peripheral CD8+ T lymphocytes in non-PD-1high expressers and PD-1high expressers. (C) Receiver operative curves describing the performance of the frequency of peripheral PD-1high CD8+ T lymphocytes in distinguishing non-PD-1high expressers and PD-1high expressers. (D) Correlation between PD-1high cells among tumor-infiltrating FoxP3–CD4+ T lymphocytes and PD-1high cells among peripheral FoxP3–CD4+ T lymphocytes. (E) Frequency of PD-1high cells among peripheral FoxP3–CD4+ T lymphocytes in non-PD-1high expressers and PD-1high expressers. (F) Receiver operating characteristic curves describing the performance of the frequency of peripheral PD-1high FoxP3–CD4+ T lymphocytes in distinguishing non-PD-1high expressers and PD-1high expressers. (G) Correlation between PD-1high cells among tumor-infiltrating FoxP3+CD4+ T lymphocytes and PD-1high cells among peripheral FoxP3+CD4+ T lymphocytes. (H) Frequency of PD-1high cells among peripheral FoxP3+CD4+ T lymphocytes in non-PD-1high expressers and PD-1high expressers. (I) Receiver operating characteristic curves describing the performance of the frequency of peripheral PD-1high FoxP3+CD4+ T lymphocytes in distinguishing non-PD-1high expressers and D-1high expressers. AUC, area under the curve; FoxP3, forkhead box protein P3; NSCLC, non-small-cell lung cancer; N.S, not significant; PD-1, programmed cell death-1; TILs, tumor-infiltrating lymphocytes. *p<0.05; **p<0.01; ****p<0.0001.
Discussion
Reinvigoration of tumor-specific T cells with PD-1 blockade has been established as a standard armamentarium for advanced NSCLC.39 However, previous studies did not address the distinct exhaustion features of TILs in the context of clinicopathological variables,33 40 largely limiting the clinical usefulness of these studies. Here, we describe the properties of TILs with differential PD-1 expression in NSCLC that correlated with the distinct features of T-cell exhaustion and activation. Furthermore, we showed that NSCLC patients exhibited profound differences in terms of T-cell exhaustion, indicating that a different response to PD-1 blockade relies on the discrete exhaustion features of TILs at an individual patient level. Finally, we demonstrated that the simultaneous targeting of immune checkpoint inhibitory receptors significantly restores T-cell functionality, assigning translational relevance to current clinical trials and treatment strategies.
The persistent antigenic stimulation witnessed in chronic viral infections and cancer ultimately causes eccentric types of T-cell dysfunction, collectively referred to as T-cell exhaustion.1–3 During the process of T-cell exhaustion, T cells undergo unique cellular and molecular processes, displaying loss of effector functions, metabolic inertia, diminished self-renewal capacity with loss of stemness, and epigenetic and transcriptional reprogramming. Upregulation of immune checkpoint inhibitory receptors such as PD-1 and CTLA-4 evidently occurs in T-cell exhaustion,9 highlighting the therapeutic potential of the pharmacological inhibition of these receptors as new strategies for redirecting exhausted T cells to combat cancers. However, an immune checkpoint blockade does not elicit a treatment response in the majority of patients,17 largely limiting its therapeutic efficacy. Notably, an intricate understanding of T-cell exhaustion using clinical samples is warranted for appropriate patient selection, with suggestions of relevant predictive biomarkers and for enhancing therapeutic efficacy through combination strategies.
Previously, Thommen et al identified PD-1 as a marker for defining transcriptionally and functionally distinct CD8+ TILs with predictive implications.33 Here, we subdivided CD8+ TILs into the PD-1neg, PD-1int, and PD-1high populations based on PD-1 expression. PD-1high CD8+ TILs showed prominent expression of exhaustion-related and activation-related markers compared with PD-1neg and PD-1int CD8+ T cells. More importantly, the enrichment pattern of PD-1high CD8+ TILs identified two NSCLC patient subgroups, and was associated with histological subtypes, PD-L1 expression, alterations in EGFR and ALK, and history of smoking based on larger cohorts with 96 patients, previously not attempted.33 Furthermore, the ex vivo reinvigoration of CD8+ TILs was successfully achieved by the combination blockade of PD-1 and CTLA-4, confirming the efficacy of the combined immune checkpoint blockade. Lastly, phenotypes of CD8+ T cells from peripheral blood were highly correlated with those from the TME, suggesting the validity of non-invasive biomarker analysis through blood sampling. Collectively, our findings underscore the translational relevance of a detailed characterization of T-cell exhaustion using clinical samples for leveraging immunotherapeutic strategies in a more efficient manner for NSCLC treatment.
Our study highlights several important issues spanning from T-cell exhaustion biology to routine clinical practice. First, exhausted T-cell populations showed the robust upregulation of markers associated with T-cell activation, indicating that T-cell exhaustion inevitably occurs in the context of T-cell activation. As demonstrated in a previous study, these PD-1high CD8+ T cells are predicted to be reactive to clonal neoantigens commonly shared by tumor cells, which has therapeutic relevance.41 Moreover, PD-1high TILs from NSCLC patients share key characteristics with exhausted T cells described in previous studies,33 34 such as coexpression of multiple immune checkpoint inhibitory receptors as well as tissue-resident markers; they also clearly express higher levels of ICOS and Ki-67 compared with PD-1int or PD-1neg TILs. A striking aspect of this phenomenon is that costimulatory signals can be reinforced for further functional restoration of exhausted T cells. The largely restricted tumor reactivity of PD-1high TILs suggested by recent studies also supports the clinical utility of the simultaneous targeting of coinhibitory and costimulatory receptors.33
Second, enrichment of PD-1high CD8+ TIL was associated with worse clinical outcomes, thus revealing that the T-cell exhaustion status can be used for the incorporation of immune checkpoint blockade in adjuvant treatment strategies. Guo et al already identified that the balance between pre-exhausted T cell subset (CD8-C4-GZMK and CD8-C5-ZNF683 clusters with lower expression of immune checkpoint receptors) and exhausted T cell subset (CD8-C6-LAYN cluster with higher expression of immune checkpoint receptors) was associated with prognosis,40 consistent with our findings regarding PD-1 expression on CD8+ TILs implicated in phenotypic and functional characteristics as well as prognosis. Currently, several studies are investigating whether adjuvant immune checkpoint blockade therapy can lower the incidence of disease relapse and improve survival (NCT02504372, NCT02595944, NCT02486718, and NCT02273375). These studies largely depend on pathological staging for patient enrollment, extrapolating previous strategies for adjuvant chemotherapy. However, our analyses reveal that the extent of T-cell exhaustion was not apparently correlated with disease stage. As in the case of the adjuvant trials focusing on molecular alterations such as EGFR mutations,42 we propose that defining the T-cell exhaustion status can be useful in the enrichment of patients who may derive clinical benefits from immune checkpoint blockade in adjuvant settings. To support this hypothesis, the frequency of PD-1high expressers in our study cohort is similar to the response rate for PD-1 blockade in the prospective studies.10 14 15 Although there are conflicting data as to whether EGFR and ALK wild-type tumor are associated with a worse prognosis after surgical resection and an improved response to checkpoint inhibitors,43–48 our study suggests that the T-cell exhaustion status can be utilized to select patient populations that would be expected to most highly benefit from adjuvant treatment with immune checkpoint inhibitors.
Third, clinicopathological variables traditionally known to be associated with the benefits of PD-1 blockade largely predicted the presence of distinct PD-1high populations in CD8+ TILs, although some degree of discordance is evident. For example, PD-L1 expression was higher in PD-1high expressers. In addition, PD-1high expressers were negatively enriched in EGFR or ALK-altered NSCLC, which is known to have a lower tumor mutation burden with corresponding lower neoantigen levels,49 50 decreased infiltration of CD8+ T cells,26 and a reduced IFN-γ signature.49 51 Smokers were also frequently observed among PD-1high expressers (figure 3B), consistent with a previous study demonstrating associations among smoking, chronic obstructive pulmonary disease, and the response to anti-PD-L1/PD-1 inhibitors.52 This finding is largely consistent with previous studies revealing the plausible association between PD-L1 expression, tumor mutation burden, smoking history, oncogenic driver mutations, and response to PD-1 blockade.20 25 26 However, lower PD-L1 expression, as well as the presence of EGFR mutations and ALK rearrangements, do not exclude the possibility of the clinical benefit of PD-1 blockade, reflecting that the cross-talk between tumor-intrinsic molecular alterations and T-cell exhaustion comprehensively dictates the response to PD-1 blockade. Opposing roles of IFN on tumor cells and immune cells can be an example to finely determine anti-PD-1 response.53 Further comprehensive explorations to define multiple factors to dictate response for PD-1 blockade is required in future studies with a larger sample size.
Fourth, our ex vivo analysis of TILs confirmed that the combined blockade of PD-1 and CTLA-4 cooperatively enhanced effector cytokine production, providing direct evidence for the enhanced efficacy of dual checkpoint inhibitors. In line with this, the CheckMate-227 (NCT02477826) and CheckMate-9LA (NCT03215706) studies have shown robust antitumor activity following the combined use of PD-1 and CTLA-4 inhibitors, which ultimately translated into improved overall survival. Based on these results, the US Food and Drug Administration has approved pembrolizumab plus ipilimumab and nivolumab plus ipilimumab with chemotherapy for the first-line treatment of NSCLC patients. Future investigations of the combined blockade of the PD-1/PD-L1 axis and other immune checkpoint coinhibitory receptors such as TIGIT (NCT03563716) would be clinically relevant. Furthermore, investigating whether inhibition of IFN signaling pathway provoked by checkpoint inhibitors in a tumor-specific manner would bring further benefit for patients treated immunotherapy would be another intriguing area of future research, based on the study conducted by Benci et al.53
Lastly, non-invasive blood-based biomarker analysis can be used to monitor the T-cell exhaustion status in the TME. Our study showed a noteworthy correlation between the frequency of the PD-1high population in CD8+ T cells from the peripheral blood and tumors. Notably, Huang et al54 demonstrated clonal overlap between CD8+ T cells from the tumor and peripheral blood, providing evidence that peripheral CD8+ T cells can be used as a non-invasive marker for the suspected characteristics of CD8+ TILs. Indeed, several studies have proposed using PD-1+CD8+ T cells or their characteristics as predictors of the response to checkpoint inhibitors.54–56 The analysis of the currently suggested biomarkers for PD-1 blockade, such as PD-L1 expression, tumor mutation burden, pre-existing T cell infiltration, and transcription signatures, necessitates the collection of tissue specimens, which is apparently associated with a higher risk of procedure-related comorbidity and mortality, especially in NSCLC patients.27 57 Considering these aspects, blood-based biomarker analysis can be a valuable tool for serial immune monitoring. We anticipate that our findings can be validated by matching tumor samples with blood samples in future clinical trials.
In conclusion, we showed a distinct exhaustion status of TILs in patients with NSCLC at both the intraindividual and interindividual levels. Our results indicate that the characteristics of T cell exhaustion measured using clinical samples have therapeutic implications in terms of understanding T-cell exhaustion biology, deducing relevant biomarkers, and designing future clinical trials, providing compelling evidence for the fact that the exhaustion features of TILs determine the response to PD-1 blockade. Future studies should examine whether the selective targeting of PD-1high populations can further enhance the clinical benefits of immunotherapy in view of the distinguished cellular states of these cells, and whether PD-1 blockade can systemically influence the T-cell fate with regard to their exhaustion in the TME.
Data availability statement
Data are available on reasonable request. Data are available from the corresponding author HRK on reasonable request.
Ethics statements
Patient consent for publication
Ethics approval
This study was approved by the Yonsei Cancer Center Institutional Review Board (4-2018-1210).
Acknowledgments
We thank all patients who participated in this study.
References
Supplementary materials
Supplementary Data
This web only file has been produced by the BMJ Publishing Group from an electronic file supplied by the author(s) and has not been edited for content.
Footnotes
Contributors Responsible for the overall content as guarantor: HRK. Conception and design: SYP, S-JH, and HRK. Financial support: CGK, SYP, S-JH, and HRK. Provision of study materials or patients: HSS, HIY, SYP, S-JH, and HRK. Collection and assembly of data: CGK, GK, KHK, SP, SS, and DY. Data analysis and interpretation: CGK, GK, KHK, SP, SS, HSS, HIY, SYP, S-JH, and HRK. Manuscript writing: CGK, GK, SYP, S-JH, and HRK. All authors read and approved the final manuscript.
Funding This research was supported by the Bio & Medical Technology Development Program of the National Research Foundation funded by the Ministry of Science and ICT (NRF-2019M3A9B6065231, 2017M3A9E802971, and 2017M3A9E9072669 to HRK and 2019M3A9B6065221, 2018R1A2A1A05076997, and 2017R1A5A1014560 to S-JH), Severance Hospital Research fund for Clinical excellence (C-2021-0016 to HRK), and Young Medical Scientist Research Grant Program of the Daewoong Foundation (DY20206P to CGK).
Competing interests None declared.
Provenance and peer review Not commissioned; externally peer reviewed.
Supplemental material This content has been supplied by the author(s). It has not been vetted by BMJ Publishing Group Limited (BMJ) and may not have been peer-reviewed. Any opinions or recommendations discussed are solely those of the author(s) and are not endorsed by BMJ. BMJ disclaims all liability and responsibility arising from any reliance placed on the content. Where the content includes any translated material, BMJ does not warrant the accuracy and reliability of the translations (including but not limited to local regulations, clinical guidelines, terminology, drug names and drug dosages), and is not responsible for any error and/or omissions arising from translation and adaptation or otherwise.