Article Text

Abstract
Background The effects of non-amplification short variant (SV) mutations in CD274 (programmed death-ligand 1 (PD-L1)) on PD-L1 protein expression and immune checkpoint inhibitors (ICPIs) therapy are unknown. Here, we present a retrospective analysis of CD274 mutations detected by comprehensive genomic profiling (CGP) and correlate these results with tumor-cell PD-L1 immunohistochemistry (IHC)-based expression assessment to better understand the relationship between mutations and protein expression of PD-L1.
Methods CGP was performed on hybridization-captured, adaptor ligation-based libraries using DNA and/or RNA extracted from 314,631 tumor samples that were sequenced for up to 406 cancer-related genes and select gene rearrangements. PD-L1 IHC was performed on a subset of cases (n=58,341) using the DAKO 22C3 PD-L1 IHC assay and scored with the tumor proportion score (TPS).
Results Overall, the prevalence of CD274 SV mutations was low (0.3%, 1081/314,631) with 577 unique variants. The most common CD274 SV mutations were R260H (n=51), R260C (n=18), R125Q (n=12), C272fs*13 (n=11), R86W (n=10), and R113H (n=10). The prevalence of CD274 mutations varied depending on tumor type with diffuse large B-cell lymphoma (1.9%, 19/997), cutaneous squamous cell carcinoma (1.6%, 14/868), endometrial adenocarcinoma (1.0%, 36/3740), unknown primary melanoma (0.9%, 33/3679), and cutaneous melanoma (0.8%, 32/3874) having the highest frequency of mutations. Of the R260H cases concurrently tested with PD-L1 IHC, most (81.8%, 9/11) had no PD-L1 expression, which contrasts to the five E237K cases where most (80%, 4/5) had PD-L1 expression. In addition, we saw a significantly lower level of PD-L1 expression in samples with a clonal truncating variant (nonsense or frameshift indel) when compared with samples with a subclonal truncating variants (mean: TPS=1 vs TPS=38; p<0.001), and also in clonal versus subclonal missense mutations (mean: TPS=11 vs TPS=22, respectively; p=0.049)
Conclusions We defined the landscape of CD274 mutations in a large cohort of tumor types that can be used as a reference for examining CD274 mutations as potential resistance biomarkers for ICPI. Furthermore, we presented novel data on the correlation of CD274 mutations and PD-L1 protein expression, providing important new information on the potential functionality of these mutations and can serve as a basis for future research.
- immunotherapy
- tumor biomarkers
Data availability statement
All data relevant to the study are included in the article or uploaded as supplementary information. The data generated by the research that support our article will be provided in the supplements.
This is an open access article distributed in accordance with the Creative Commons Attribution Non Commercial (CC BY-NC 4.0) license, which permits others to distribute, remix, adapt, build upon this work non-commercially, and license their derivative works on different terms, provided the original work is properly cited, appropriate credit is given, any changes made indicated, and the use is non-commercial. See http://creativecommons.org/licenses/by-nc/4.0/.
Statistics from Altmetric.com
Introduction
Immune checkpoint inhibitors (ICPIs) that block the programmed death-ligand 1 and programmed cell death protein 1 (PD-L1/PD-1) axis have shown great clinical utility in a wide variety of solid tumors and hematologic malignancies.1–4 Multiple companion diagnostics (CDx) for ICPI have been developed and subsequently approved by the United States Food and Drug Administration.5 6 A frequent ICPI CDx utilized for multiple tumor types is PD-L1 immunohistochemistry (IHC) which can detect PD-L1 protein expression/overexpression on tumor cells and tumor-infiltrating immune cells. Multiple clinical trials have shown that in specific tumor types, a certain level of PD-L1 protein expression is necessary in the tumor microenvironment for a PD-L1/PD-1 inhibitor to be efficacious.4 7 8
The PD-L1 protein is encoded by the approximately 17.6 kb CD274 (PD-L1) gene located on chromosome 9p24.1.9 The CD274 Matched Annotation from NCBI and EMBL-EBI transcript (ENST00000381577.4) encodes for a type 1 transmembrane protein that is 290 amino acids long and has immunoglobulin V-like and C-like domains.10 Currently, in large public genomic databases like COSMIC, only 229 CD274 non-amplification short variant (SV)-mutated samples have been reported.11 12
Previously, two large studies examined PD-L1 protein expression in a variety of tumor types; however, the published literature contains limited data on CD274 SV mutations.13 14 Here, we present the landscape of CD274 SV mutations detected by comprehensive genomic profiling (CGP) in a large pan-cancer genomic database. In addition, for a subset of these SV mutations, we evaluated PD-L1 protein expression via IHC to better understand the effect of these mutations on protein expression of PD-L1.
Materials and methods
Sample cohort
We analyzed all cases that underwent CGP testing at Foundation Medicine, Inc between January 2014 and August 2020. Formalin-fixed, paraffin-embedded (FFPE) tissue of either whole section samples, biopsies, or cytology specimens were received as paraffin blocks or unstained slides from outside institutions during routine clinical care. A board-certified pathologist assigned a diagnosis for each specimen based on microscopic examination of a H&E stained slide from the FFPE tissue, the accompanying pathology report, and additional information provided by the ordering physician.
Comprehensive genomic Profiling
CGP was performed on hybridization-captured, adaptor ligation-based libraries using DNA and/or RNA extracted from FFPE tumor in a Clinical Laboratory Improvement Amendments (CLIA)-certified and College of American Pathologists (CAP)-accredited laboratory (Foundation Medicine, Inc, Cambridge, Massachusetts, USA). The samples were sequenced for up to 406 cancer-related genes and select gene rearrangements.15 CD274 non-amplification SV mutations were defined as missense mutations, truncations, splice site mutations, and insertion/deletions, as previously described.15 CD274 amplification was defined as ploidy +4. Tumor mutational burden (TMB) was determined on up to 1.24 Mb of sequenced DNA and TMB ≥10 mutations/Mb (mut/Mb) was considered TMB-High per CDx approval.16 17 Microsatellite instability (MSI) was performed from DNA sequencing up to 114 loci and MSI-High (MSI-H) was considered positive per CDx approval.18 19 In addition, as research use only (RUO), ultraviolet mutational signatures were called as described by Zehir et al.20
Clonality, predicted germline versus predicted somatic mutation, and predicted missense functionality
For the purposes of this study, a subclonal SV mutation was defined as a sample where <50% of tumor cells were predicted to harbor the variant based on both the variant allele fraction and the pathologic and/or computational tumor cell purity estimates. A RUO somatic germline zygosity (SGZ) bioinformatics algorithm was used to determine whether mutations were likely somatic or germline, as previously described.21 We assessed CD274 missense mutation’s functionality with several in silico methods including SIFT, MutationTaster, fathmm-MKL, and MetaSVM and recalibrated the scores to a rankscore so they can be compared with each other.22–25 The rankscore was on a scale of 0 to 1 with 0 being predicted to be a non-functional protein and 1 being predicted to be a functional protein.
DAKO PD-L1 IHC 22C3 assay
For a subset of cases, the PD-L1 DAKO 22C3 assay was run according to manufacturer instructions in a CLIA-certified and CAP-accredited laboratory (Foundation Medicine, Inc, Morrisville, North Carolina, USA).26 The IHC cases were interpreted by board-certified pathologists specifically trained on the DAKO tumor proportion score (TPS) method, where tumor cell expression of PD-L1 was quantified. The DAKO TPS scoring method was defined as TPS=# PD-L1 positive tumor cells/(total # of PD-L1 positive + PD-L1 negative tumor cells).27
Results
Landscape of CD274 SV mutations
Overall, the frequency of CD274 SV mutations was low (0.3%, 1081/314,631) in our cohort of 314,631 samples. A total of 577 unique variants were discovered; some mutations were recurrent, while others occurred only once in the entire cohort (table 1). Of the 1081 mutations, 49.9% (539/1081) were from metastatic specimens and 42.3% (457/1081) were from primary specimens. In 7.8% (85/1081) of cases it was unknown whether it was a primary or metastatic specimen. Of the 1081 samples with CD274 SV mutations, only 1.4% (15/1081) had cooccurring CD274 amplification.
The most common CD274 mutations were R260H (n=51), R260C (n=18), R125Q (n=12), C272fs*13 (n=11), R86W (n=10), and R113H (n=10) (table 1, figure 1). R260C/H was the most frequent recurrent missense mutation; both substitutions have been observed in the germline of healthy subjects (gnomAD). Based on the SGZ algorithm, we found that 29.0% (20/69) of the codon R260 mutations were likely somatic and 55.1% (38/69) were likely germline. The algorithm could not predict whether the variant was germline or somatic in 16.0% (11/69) of the samples. In addition, when we examined all the missense mutations (n=974), we saw that 51.0% (497/974) were predicted to be somatic, 24.7% (241/974) were predicted to be germline, and the algorithm could not predict whether the variant was germline or somatic in 24.2% (236/974) of the cases (online supplemental table 1). In addition, C272fs*13 is at an indel at a poly-A homopolymer, a sequence context that is highly mutable in the setting of MSI-H status. This mutation was significantly enriched in the MSI-H group (0.01%, 5/5139) when compared with the non-MSI-H group (0.002%, 6/309,492) (Fisher’s exact test, p<0.0001), suggesting that the variant is often a result of mismatch repair protein deficiency. This finding is reflected in the high CD274 SV mutation frequency in non-serous endometrial adenocarcinomas in this study.
Supplemental material
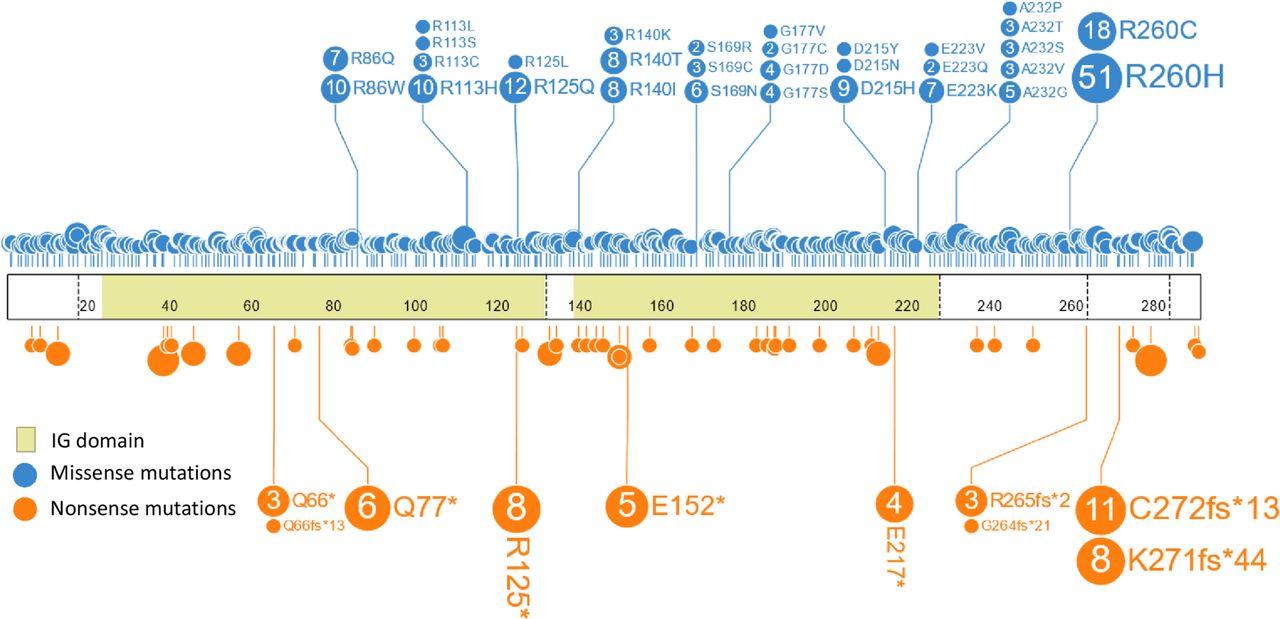
Lollipop plot of all the missense and nonsense mutations in the cohort. The most common CD274 mutations were R260H (n=51), R260C (n=18), R125Q (n=12), C272fs*13 (n=11), R86W (n=10), and R113H (n=10).
The types of mutations in this cohort also varied, with missense mutations being the most common (83.8%, 906/1081) and insertion/deletions being less common (0.8%, 9/1081) (table 2). Multiple samples had complex CD274 mutations, defined as more than one CD274 genomic alteration observed in the sample. The most common type of complex CD274 mutation was alterations with two missense mutations (1.9%, 21/1081), while other complex mutations consisted of a CD274 mutation with concurrent CD274 amplification (1.4%, 15/1081) and/or rearrangement (0.3%, 3/1081).
The prevalence of CD274 mutations also varied depending on tumor type. The top five tumor types (minimum 800 total samples) with the highest rates of CD274 mutations in descending order were: diffuse large B-cell lymphoma (1.9%, 19/997), cutaneous squamous cell carcinoma (1.6%, 14/868), endometrial adenocarcinoma (1.0%, 36/3740), unknown primary melanoma (0.9%, 33/3679), and cutaneous melanoma (0.8%, 32/3874) (figure 2, (online supplemental table 2). Interestingly, three of the five tumor types with the highest prevalence of CD274 mutations usually occur on the skin. When we examined the mean TMB for these cases, we saw very high mean TMB and median TMB (cutaneous squamous cell carcinoma (151 mut/Mb, 100 mut/Mb), cutaneous melanoma (133 mut/Mb, 126 mut/Mb), unknown primary melanoma (125 mut/Mb, 92 mut/Mb), respectively), suggesting that these were likely caused by ultraviolet exposure-induced hypermutation. This was further supported by the high prevalence of ultraviolet mutational signature in these tumor types (cutaneous squamous cell carcinoma (84.6%, 11/13), cutaneous melanoma (93.8%, 30/32), and unknown primary melanoma (100%, 32/32). Whether these CD274 mutations are random ‘passenger’ mutations due to the high mutation rate should be further investigated.

Longtail plot showing the prevalence of CD274 mutations in different tumor types. Tumor types (at least 800 total samples) with the highest rates of CD274 mutations in descending order were: diffuse large B-cell lymphoma, cutaneous squamous cell carcinoma, uterine endometrial adenocarcinoma, unknown primary melanoma, and cutaneous melanoma.
Correlation of CD274 mutations with PD-L1 IHC tumor cell expression and predicted functionality models
Of the 1081 cases with CD274 mutations, 19.7% (213/1081) cases had PD-L1 IHC data; and of the 313,550 cases without CD274 mutations, 18.6% (58,218/313,550) cases had PD-L1 IHC data.
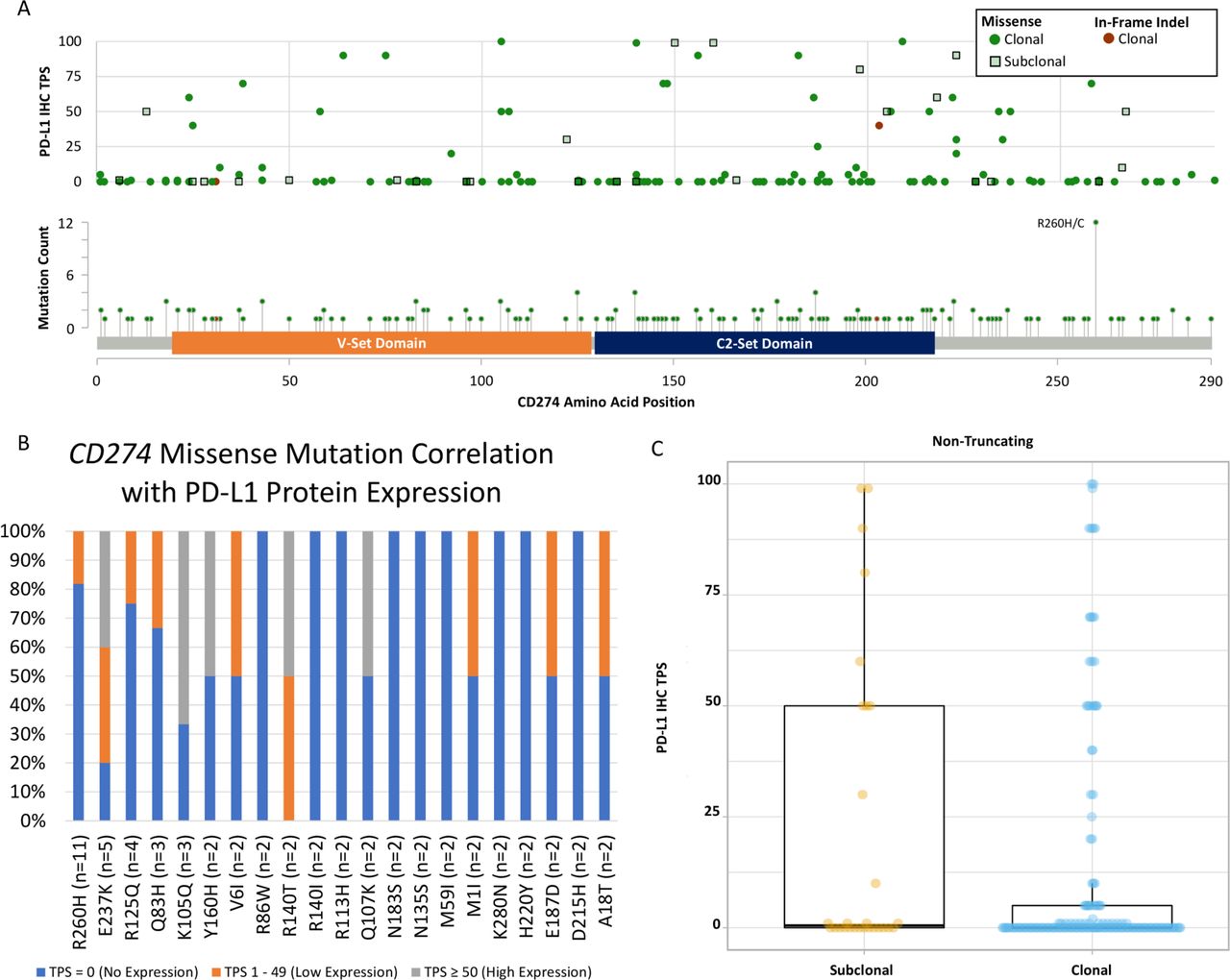
Most of the CD274 non-truncating mutations had low to no tumor-cell expression of PD-L1 (figure 3A, online supplemental table 3). Of the 11 R260H cases concurrently tested with PD-L1 IHC, 81.8% (9/11) had no PD-L1 expression and 2 (18.2%) cases had low PD-L1 expression. Of the 5 E237K cases, 20% (1/5) had no PD-L1 expression, 40% (2/5) had low PD-L1 expression, and 40% (2/5) had high PD-L1 expression (figure 3B). This difference in protein expression of the two mutations was significantly different (Fisher’s exact test, p=0.036). When we compared the PD-L1 protein expression of cases with CD274 missense mutations (n=153) and cases without CD274 mutations (n=58 218), we saw a lower level of PD-L1 IHC expression in the cases with CD274 mutations, though the difference was not statistically significant (mean: TPS=11 vs 13, respectively; analysis of variance (ANOVA), p=0.404). When examining the clonal (n=153) versus subclonal (n=28) missense mutations, we saw significantly lower PD-L1 IHC expression in the clonal missense mutation (mean: TPS=11 vs 22, respectively; ANOVA, p=0.049) (figure 3C).

Correlation of CD274 non-truncating mutations with PD-L1 IHC tumor-cell expression. (A) Among non-truncating variants, 181 samples with missense substitutions and two in-frame indels were identified (lower). A subset of the variants was recurrent, with 12 samples harboring a substitution at R260. PD-L1 TPS scores corresponding to each sample with a non-truncating variant shown (upper). Subclonal variants are denoted with squares. (B) Correlation of CD274 missense mutations and PD-L1 protein expression in mutations where at least two cases with PD-L1 IHC performed. (C) When examining the clonal (n=153) versus subclonal (n=28) missense mutations, we saw significantly lower PD-L1 IHC expression in the clonal missense mutation (mean: TPS=11 vs 22, respectively; ANOVA, p=0.049). Note: Subclonal variants were defined as samples where <50% of tumor cells were predicted to harbor the variant based on the variant allele fraction and the pathologic and/or computational tumor-cell purity estimates. ANOVA, analysis of variance; IHC, immunohistochemistry; PD-L1, programmed cell death-ligand 1; TPS, tumor proportion score.
In addition, we examined the predicted functionality of each CD274 missense mutation with multiple functionality prediction models including SIFT, MutationTaster, fathmm-MKL, and MetaSVM (online supplemental table 4). However, the predicted functionality did not have any significant correlation with PD-L1 IHC expression (online supplemental figure 1).
Thirty-nine putative truncating variants with concurrent PD-L1 IHC testing were identified, including 12 nonsense mutations, 10 frameshift indels, and 7 canonical splice variants (figure 4A, online supplemental table 2). When we compared the PD-L1 protein expression of cases with CD274 clonal truncating variant (nonsense or frameshift indel) (n=18) and cases without CD274 mutations (n=58,218), we saw a lower level of PD-L1 expression in the cases with CD274 mutations (mean: TPS=1 vs 13, respectively; ANOVA, p=0.069). In addition, we saw a significantly lower level of PD-L1 expression in samples with a clonal truncating variant (nonsense or frameshift indel) when compared with samples with subclonal truncating variants (mean: TPS=1 vs TPS=38; ANOVA, p<0.001) (figure 4B).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Correlation of CD274 truncating mutations with PD-L1 IHC tumor-cell expression. (A) Thirty-nine putative truncating variants were identified, including 12 nonsense mutations, 10 frameshift indels, and 7 canonical splice variants (lower). PD-L1 TPS scores corresponding to each sample with a non-truncating variant shown (upper). Subclonal variants are denoted with squares. (B) We saw a significantly lower level of PD-L1 expression in samples with a clonal truncating variant (nonsense or frameshift indel) when compared with samples with subclonal truncating variants (mean: TPS=1 vs TPS=38; ANOVA, p<0.001). Note: subclonal variants were defined as samples where <50% of tumor cells were predicted to harbor the variant based on the variant allele fraction and the pathologic and/or computational tumor cell purity estimates. ANOVA, analysis of variance; IHC, immunohistochemistry; PD-L1, programmed cell death-ligand 1; TPS, tumor proportion score.
Discussion
This study describes a large cohort of 1081 clinically advanced malignancies with CD274 non-amplification SV mutations, including 213 samples with concurrent PD-L1 protein expression levels. Similar to the COSMIC database (0.4%, 229/51,849), the overall prevalence across tumor types was low (0.3%, 1,081/314,631). Most of the SV mutations found were missense substitutions, with rarer nonsense and indel alterations.12 Last, the prevalence of CD274 SV mutations was higher in patients with MSI-H-associated endometrial and ultraviolet light exposed cutaneous cancers.
While most of the non-truncating SV CD274 mutations had low to no tumor-cell expression of PD-L1, the expression levels of PD-L1 differed among the various CD274 SV mutation categories. Interestingly, of the 11 R260H cases concurrently tested with PD-L1 IHC, most showed little to no PD-L1 expression, suggesting that patients with R260H mutations might not be treated with ICPI if only tested with PD-L1 IHC. The exact mechanism that causes the R260H to turn off PD-L1 expression and/or to potentially interfere with the binding of ICPI to PD-L1 ligand should be further studied, especially given the relatively high prevalence of R260H mutation in our cohort (4.7%, 51/1081). This contrasted with the five E237K cases, where most cases had some PD-L1 expression, suggesting that the E237K has little to no effect on PD-L1 protein expression. Most other variants were only observed in a single sample (with or without concurrent PD-L1 expression data) due the rarity of the mutations. This rarity of individual variants, even in a very large database of cases with CGP, makes the clinical significance of these uncommon mutations less clear. However, as more studies evaluate the PD-L1 protein expression of tumors with CD274 SV mutations, these data can be aggregated to better understand the functionality of CD274 mutations on PD-L1 protein expression.
As expected, samples with a clonal truncating mutation had a lower level of PD-L1 expression when compared with cases without a CD274 mutation and to samples with a subclonal truncating mutation. These data suggest that when a clonal truncating event is observed in CD274, PD-L1 expression is inhibited, but in the setting of a subclonal truncation, clinical assessment of sample-level PD-L1 expression is often not affected. We hypothesize that clonal truncating variants can potentially act as resistance biomarkers for ICPI due the lack of PD-L1 protein present on the tumor cells as exemplified by our PD-L1 IHC expression data. With decreased/no ligand for the PD-L1/PD-1 inhibitors to bind to, the efficacy of ICPI would likely be diminished.
In terms of CD274 missense mutations, these could mediate resistance to ICPI due to potential steric or affinity-altering interferences in the binding of the PD-L1 ligand to the PD-1 receptor, similar to a resistance mechanism described for ROS1, though further studies are needed to evaluate this hypothesis.28 29 In our cohort of cases with CD274 missense mutations, we observed a slightly lower level of PD-L1 IHC staining in the cases with CD274 missense mutations when compared with cases without CD274 mutations, and we also saw significantly lower PD-L1 IHC staining in the clonal missense mutation when compared with the subclonal missense mutations. We hypothesize that this is likely due to the lower rates of PD-L1 antibody binding (from the IHC assay) to the PD-L1 ligand on the tumor cells instead of actual lower PD-L1 protein expression. In addition, we saw no correlation between the predicted functional status of these missense mutations and PD-L1 IHC expression since these prediction algorithms do not formally consider the steric or binding affinity interferences that can result from missense mutations into the functionality assessment. Taken together, we have some preliminary data that support our hypothesis of the potential mechanism of resistance of tumors with CD274 missense mutations to ICPI, and this should be further investigated in both in vitro and clinical studies.
This study has two primary limitations. First, we only have PD-L1 IHC on a limited subset of the samples with CD274 mutations. The second limitation of this study is that we do not have any clinical outcome data of patients with CD274 mutations and treated ICPI to assess the clinical impact of ICPI on patients with CD274 mutations.
Conclusions
We presented the landscape of CD274 mutations in a large pan-cancer cohort that can be used as a reference for examining CD274 mutations as potential resistance biomarkers for ICPI. Furthermore, we presented novel data on the correlation of CD274 mutations and PD-L1 protein expression, providing important data on the potential functionality of these mutations that will serve as a basis for future research.
Data availability statement
All data relevant to the study are included in the article or uploaded as supplementary information. The data generated by the research that support our article will be provided in the supplements.
Ethics statements
Ethics approval
Approval for this study was obtained from the Western Institutional Review Board Protocol No 20152817.
References
Supplementary materials
Supplementary Data
This web only file has been produced by the BMJ Publishing Group from an electronic file supplied by the author(s) and has not been edited for content.
Footnotes
Contributors RSPH, BD and JSR were invovled in study conception/design. Provision of study material or patients were done by RSPH, BD, KM, MH, DAM, SR and JSR. Collection and/or assembly of data were performed by RH and BD. Data analysis and interpretation and manuscript writing were done by all the authors. All authors gave final approval of manuscript.
Funding The authors have not declared a specific grant for this research from any funding agency in the public, commercial or not-for-profit sectors.
Competing interests All authors of the manuscript are employees of Foundation Medicine, Inc, which is a wholly owned subsidiary of Roche and receives stock from Roche.
Provenance and peer review Not commissioned; externally peer reviewed.
Supplemental material This content has been supplied by the author(s). It has not been vetted by BMJ Publishing Group Limited (BMJ) and may not have been peer-reviewed. Any opinions or recommendations discussed are solely those of the author(s) and are not endorsed by BMJ. BMJ disclaims all liability and responsibility arising from any reliance placed on the content. Where the content includes any translated material, BMJ does not warrant the accuracy and reliability of the translations (including but not limited to local regulations, clinical guidelines, terminology, drug names and drug dosages), and is not responsible for any error and/or omissions arising from translation and adaptation or otherwise.