
Abstract
Treatments that target immune checkpoints, such as the one mediated by programmed cell death protein 1 (PD-1) and its ligand PD-L1, have been approved for treating human cancers with durable clinical benefit1,2. However, many patients with cancer fail to respond to compounds that target the PD-1 and PD-L1 interaction, and the underlying mechanism(s) is not well understood3,4,5. Recent studies revealed that response to PD-1–PD-L1 blockade might correlate with PD-L1 expression levels in tumour cells6,7. Hence, it is important to understand the mechanistic pathways that control PD-L1 protein expression and stability, which can offer a molecular basis to improve the clinical response rate and efficacy of PD-1–PD-L1 blockade in patients with cancer. Here we show that PD-L1 protein abundance is regulated by cyclin D–CDK4 and the cullin 3–SPOP E3 ligase via proteasome-mediated degradation. Inhibition of CDK4 and CDK6 (hereafter CDK4/6) in vivo increases PD-L1 protein levels by impeding cyclin D–CDK4-mediated phosphorylation of speckle-type POZ protein (SPOP) and thereby promoting SPOP degradation by the anaphase-promoting complex activator FZR1. Loss-of-function mutations in SPOP compromise ubiquitination-mediated PD-L1 degradation, leading to increased PD-L1 levels and reduced numbers of tumour-infiltrating lymphocytes in mouse tumours and in primary human prostate cancer specimens. Notably, combining CDK4/6 inhibitor treatment with anti-PD-1 immunotherapy enhances tumour regression and markedly improves overall survival rates in mouse tumour models. Our study uncovers a novel molecular mechanism for regulating PD-L1 protein stability by a cell cycle kinase and reveals the potential for using combination treatment with CDK4/6 inhibitors and PD-1–PD-L1 immune checkpoint blockade to enhance therapeutic efficacy for human cancers.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 51 print issues and online access
$199.00 per year
only $3.90 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout




Similar content being viewed by others

References
Zou, W., Wolchok, J. D. & Chen, L. PD-L1 (B7-H1) and PD-1 pathway blockade for cancer therapy: Mechanisms, response biomarkers, and combinations. Sci. Transl. Med. 8, 328rv4 (2016)
Boussiotis, V. A. Molecular and biochemical aspects of the PD-1 checkpoint pathway. N. Engl. J. Med. 375, 1767–1778 (2016)
Gotwals, P. et al. Prospects for combining targeted and conventional cancer therapy with immunotherapy. Nat. Rev. Cancer 17, 286–301 (2017)
Sharma, P. & Allison, J. P. The future of immune checkpoint therapy. Science 348, 56–61 (2015)
Mahoney, K. M., Rennert, P. D. & Freeman, G. J. Combination cancer immunotherapy and new immunomodulatory targets. Nat. Rev. Drug Discov. 14, 561–584 (2015)
Herbst, R. S. et al. Predictive correlates of response to the anti-PD-L1 antibody MPDL3280A in cancer patients. Nature 515, 563–567 (2014)
Iwai, Y. et al. Involvement of PD-L1 on tumor cells in the escape from host immune system and tumor immunotherapy by PD-L1 blockade. Proc. Natl Acad. Sci. USA 99, 12293–12297 (2002)
Otto, T. & Sicinski, P. Cell cycle proteins as promising targets in cancer therapy. Nat. Rev. Cancer 17, 93–115 (2017)
Casey, S. C. et al. MYC regulates the antitumor immune response through CD47 and PD-L1. Science 352, 227–231 (2016)
Dorand, R. D. et al. Cdk5 disruption attenuates tumor PD-L1 expression and promotes antitumor immunity. Science 353, 399–403 (2016)
Li, C. W. et al. Glycosylation and stabilization of programmed death ligand-1 suppresses T-cell activity. Nat. Commun. 7, 12632 (2016)
Lim, S. O. et al. Deubiquitination and stabilization of PD-L1 by CSN5. Cancer Cell 30, 925–939 (2016)
Schiff, P. B. & Horwitz, S. B. Taxol stabilizes microtubules in mouse fibroblast cells. Proc. Natl Acad. Sci. USA 77, 1561–1565 (1980)
Malumbres, M. & Barbacid, M. Mammalian cyclin-dependent kinases. Trends Biochem. Sci. 30, 630–641 (2005)
Hydbring, P., Malumbres, M. & Sicinski, P. Non-canonical functions of cell cycle cyclins and cyclin-dependent kinases. Nat. Rev. Mol. Cell Biol. 17, 280–292 (2016)
Bates, S. et al. CDK6 (PLSTIRE) and CDK4 (PSK-J3) are a distinct subset of the cyclin-dependent kinases that associate with cyclin D1. Oncogene 9, 71–79 (1994)
Lees, E ., Faha, B ., Dulic, V ., Reed, S. I. & Harlow, E. Cyclin E/cdk2 and cyclin A/cdk2 kinases associate with p107 and E2F in a temporally distinct manner. Genes Dev. 6, 1874–1885 (1992)
Takaki, T. et al. Preferences for phosphorylation sites in the retinoblastoma protein of D-type cyclin-dependent kinases, Cdk4 and Cdk6, in vitro. J. Biochem. 137, 381–386 (2005)
Fry, D. W. et al. Specific inhibition of cyclin-dependent kinase 4/6 by PD 0332991 and associated antitumor activity in human tumor xenografts. Mol. Cancer Ther. 3, 1427–1438 (2004)
Parry, D., Bates, S., Mann, D. J. & Peters, G. Lack of cyclin D–Cdk complexes in Rb-negative cells correlates with high levels of p16INK4/MTS1 tumour suppressor gene product. EMBO J. 14, 503–511 (1995)
Lukas, J. et al. Retinoblastoma-protein-dependent cell-cycle inhibition by the tumour suppressor p16. Nature 375, 503–506 (1995)
Soucy, T. A. et al. An inhibitor of NEDD8-activating enzyme as a new approach to treat cancer. Nature 458, 732–736 (2009)
Genschik, P., Sumara, I. & Lechner, E. The emerging family of CULLIN3-RING ubiquitin ligases (CRL3s): cellular functions and disease implications. EMBO J. 32, 2307–2320 (2013)
Barbieri, C. E. et al. Exome sequencing identifies recurrent SPOP, FOXA1 and MED12 mutations in prostate cancer. Nat. Genet. 44, 685–689 (2012)
Cancer Genome Atlas Research. The molecular taxonomy of primary prostate cancer. Cell 163, 1011–1025 (2015)
Sato, Y. et al. Integrated molecular analysis of clear-cell renal cell carcinoma. Nat. Genet. 45, 860–867 (2013)
Xu, J. et al. Genome-wide association study in Chinese men identifies two new prostate cancer risk loci at 9q31.2 and 19q13.4. Nat. Genet. 44, 1231–1235 (2012)
Gan, W. et al. SPOP promotes ubiquitination and degradation of the ERG oncoprotein to suppress prostate cancer progression. Mol. Cell 59, 917–930 (2015)
da Fonseca, P. C. et al. Structures of APC/CCdh1 with substrates identify Cdh1 and Apc10 as the D-box co-receptor. Nature 470, 274–278 (2011)
Goel, S. et al. CDK4/6 inhibition triggers anti-tumour immunity. Nature 548, 471–475 (2017)
Dai, X. et al. Prostate cancer-associated SPOP mutations confer resistance to BET inhibitors through stabilization of BRD4. Nat. Med. 23, 1063–1071 (2017)
Wan, L. et al. The APC/C E3 ligase complex activator FZR1 restricts BRAF oncogenic function. Cancer Discov. 7, 424–441 (2017)
Gao, D. et al. Phosphorylation by Akt1 promotes cytoplasmic localization of Skp2 and impairs APCCdh1-mediated Skp2 destruction. Nat. Cell Biol. 11, 397–408 (2009)
Yu, Q. et al. Requirement for CDK4 kinase function in breast cancer. Cancer Cell 9, 23–32 (2006)
Sicinska, E. et al. Requirement for cyclin D3 in lymphocyte development and T cell leukemias. Cancer Cell 4, 451–461 (2003)
Hamanishi, J. et al. Programmed cell death 1 ligand 1 and tumor-infiltrating CD8+ T lymphocytes are prognostic factors of human ovarian cancer. Proc. Natl Acad. Sci. USA 104, 3360–3365 (2007)
Wang, H. et al. The metabolic function of cyclin D3-CDK6 kinase in cancer cell survival. Nature 546, 426–430 (2017)
Wan, L. et al. APCCdc20 suppresses apoptosis through targeting Bim for ubiquitination and destruction. Dev. Cell 29, 377–391 (2014)
Kozar, K. et al. Mouse development and cell proliferation in the absence of D-cyclins. Cell 118, 477–491 (2004)
Choi, Y. J. et al. The requirement for cyclin D function in tumor maintenance. Cancer Cell 22, 438–451 (2012)
Sicinski, P. et al. Cyclin D1 provides a link between development and oncogenesis in the retina and breast. Cell 82, 621–630 (1995)
Yu, Q. et al. Specific protection against breast cancers by cyclin D1 ablation. Nature 411, 1017–1021 (2001)
Acknowledgements
We thank members of the Wei, Freeman, Sicinski, and Pandolfi laboratories for discussions. J.Z. is supported by the career transition award (1K99CA212292-01). W.W. is a Leukemia & Lymphoma Society research scholar. This work was supported in part by the National Institutes of Health (NIH) grants GM094777 and CA177910 (to W.W.), P01 CA080111, R01 CA202634 and R01 CA132740 (to P.S.), and P50CA101942 (to G.J.F).
Author information
Authors and Affiliations
Contributions
J.Z., X.B. and H.W. performed most of the experiments with assistance from Y.Z., Y.G., N.T.N., Y.T., Y.C., F.W., X.D., J.G., Y.H., C.F., S.R. and Y.S. Y.Z., S.R., and Y.S. performed immunohistochemistry for human prostate cancer samples. Y.G., Y.T. and Y.C. helped with mice work. J.Z., X.B., H.W., G.J.F., P.S. and W.W. designed the experiments. G.J.F., P.S. and W.W. supervised the study. J.Z. and W.W. wrote the manuscript with help from X.B., H.W., P.S. and G.J.F. All authors commented on the manuscript.
Corresponding authors
Ethics declarations
Competing interests
G.J.F. has patents and pending royalties on the PD-1 pathway from Roche, Merck, Bristol-Myers-Squibb, EMD-Serono, Boehringer-Ingelheim, AstraZeneca, DAKO and Novartis. G.J.F. has served on advisory boards for CoStim, Novartis, Roche, Eli Lilly, Bristol-Myers-Squibb, Seattle Genetics, Bethyl Laboratories, Xios, and Quiet. P.S. is a consultant and a recipient of a research grant from Novartis. No potential conflicts of interests were disclosed by other authors.
Additional information
Reviewer Information Nature thanks J. Bartek, C. Klebanoff and S. Ogawa for their contribution to the peer review of this work.
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data figures and tables
Extended Data Figure 1 PD-L1 fluctuates during cell cycle progression.
a, b, Immunoblot of WCL derived from MDA-MB-231 (a) or HCC1954 (b) cells synchronized in M phase by nocodazole treatment before release back into the cell cycle for the indicated times. c, d, Quantitative reverse transcription PCR (qRT–PCR) analyses of relative mRNA levels of PD-L1 (also known as CD274) and GAPDH from samples derived from HeLa cells synchronized in M phase by nocodazole treatment before release back to the cell cycle for the indicated times. e, Immunoblot of WCL derived from untreated HeLa cells or HeLa cells pre-treated with IFNγ (10 ng ml−1) for 12 h and then synchronized in M phase by nocodazole treatment before release back into the cell cycle for the indicated times. f, Immunoblot of WCL derived from HeLa cells stably expressing wild-type HA–MYC, or HA–MYC(T58A/S62A). Empty vector was used as a negative control. g, Immunoblot of WCL derived from wild-type HeLa cells or HeLa cells stably expressing HA–MYC; cells were synchronized in M phase by nocodazole treatment before release back into the cell cycle for the indicated times. h–k, Immunoblot of WCL derived from MC38, CT26, 4T1 or B16-F10 mouse tumour cells treated with the indicated concentration of nocodazole for 20 h before collecting. l–m, Immunoblot of WCL derived from 4T1 or CT26 mouse tumour cells treated with the indicated concentration of taxol for 20 h before collecting.
Extended Data Figure 2 Cyclin D–CDK4 negatively regulates PD-L1 protein stability.
a, b, Immunoblot analysis of WCL derived from wild-type and Ccna1−/−Ccna2−/− (a) or Ccne1−/−Ccne2−/− (b) MEFs. c, qRT–PCR analysis of relative PD-L1 mRNA levels from wild-type and Ccnd1−/−Ccnd2−/−Ccnd3−/− MEFs. Data are mean ± s.d., n = 5. NS, not significant, two-tailed t-test. d, Cell cycle profiles for wild-type and Ccnd1−/−Ccnd2−/−Ccnd3−/− MEFs, which were labelled with BrdU and propidium iodide and analysed by FACS. e, Immunoblot analysis of WCL derived from Ccnd1fl/flCcnd2−/−Ccnd3fl/fl MEFs upon expression of Cre resulting in ablation of Ccnd1 and Ccnd3. Cre was delivered via viral infection with pLenti-Cre (pLenti-EGFP used as a negative control). Cells were selected with puromycin (1 μg ml−1) for 72 h before collecting. f, Immunoblot analysis of WCL derived from Ccnd1−/−Ccnd2−/−Ccnd3−/− MEFs with stable reexpression of Ccnd1, Ccnd2 or Ccnd3. Empty vector was used as a negative control. g, Immunoblot analysis of WCL derived from mouse mammary tumours induced by MMTV-Myc in Ccnd1+/+ or Ccnd1−/− females. n = 5 mice per experimental group. h, Immunoblot analysis of WCL derived from wild-type and Cdk6−/− MEFs. i, j, Immunoblot analysis of WCL derived from MDA-MB-231 cells stably expressing shCDK6 or shCDK2 as well as shScramble as a negative control. k, l, Immunoblot analysis of WCL derived from MDA-MB-231 cells transfected with indicated constructs (k), the intensity of PD-L1 band was quantified by the ImageJ software (l). m, Immunoblot analysis of WCL derived from RB1 depleted MDA-MB-231 cells (with shScramble as a negative control) treated with palbociclib where indicated. n, o, Immunoblot analysis of WCL derived from mouse CT26 or 4T1 tumour cell lines treated with or without palbociclib or ribociclib, respectively. p, q, Immunoblot analysis of WCL derived from MDA-MB-231 cells pre-treated with palbociclib (1 μM) for 36 h before treatment with cycloheximide (CHX) for the indicated time (p) and PD-L1 protein abundance was quantified by ImageJ as indicated (q). r, Immunoblot analysis of WCL derived from 19 different cancer cell lines with indicated antibodies. s–u, Immunoblot analysis of WCL derived from MCF7 (s), T47D (t) or HLF (u) stably expressing p16 as well as empty vector as a negative control. v–x, Immunoblot analysis of WCL derived from MDA-MB-436 (v), BT549 (w) or HCC1937 (x) stably expressing three independent shRNAs targeting p16 (also known as Cdkn2a) as well as shScramble as a negative control.
Extended Data Figure 3 Treatment with the CDK4/6 inhibitor palbociclib increases PD-L1 levels in vivo.
a, b, Immunoblot analysis of WCL derived from MC38 or B16-F10 implanted tumours, in mice treated with palbociclib (150 mg kg−1 body weight, by gastric gavage) or vehicle for 7 days. n = 5 mice per experimental group. c, FACS analysis for PD-L1 or CD3+ T cell populations from B16-F10-implanted tumours in mice treated with vehicle or palbociclib for 7 days. n = 5 mice per experimental group. d, Immunoblot analysis of WCL derived from the indicated organs in mice treated with palbociclib (150 mg kg−1 body weight, by gastric gavage) or vehicle for 7 days. n = 5 mice per experimental group. e, Quantification of PD-L1 protein bands intensity in d using ImageJ. n = 5 mice per experimental group. f, Immunoblot analysis of WCL derived from 15 different organs dissected from mice with or without palbociclib treatment and MMTV-c-Myc induced breast tumours. g, Quantification of PD-L1 protein bands intensity in f using ImageJ. n = 3 biological replicates. h, In vitro kinase assay using immunoprecipitated cyclin D1–CDK4 kinase complex from livers or brains by anti-CDK4 antibody and recombinant Rb as a substrate. Note that cyclin D1–CDK4 complex in non-dividing organs (livers and brains) displayed kinase activity, which might explain why the CDK4/6 inhibitor increased PD-L1 in these organs. Data are mean ± s.d. *P < 0.05, **P < 0.01, ***P < 0.001,. two-tailed t-test.
Extended Data Figure 4 Cullin 3–SPOP promotes PD-L1 ubiquitination and subsequent degradation largely through interaction with the cytoplasmic tail of PD-L1.
a, A schematic illustration of PD-L1 with N-terminal signal peptide, extracellular domain, transmembrane domain, cytoplasmic tail and the potential SPOP-binding motif in PD-L1. b, d, Immunoblot analysis of WCL and glutathione S-transferase (GST) pull-down precipitates derived from 293T cells transfected with indicated constructs and treated with MG132 (10 μM) for 12 h before collecting. c, Immunoblot analysis of WCL derived from PC3 stably expressing shCullin 3. e, g, Immunoblot analysis of WCL and immunoprecipitate derived from HEK293T cells transfected with indicated constructs and treated with MG132 (10 μM) for 12 h before collecting. f, Immunoblot of WCL and Ni–NTA pull-down products derived from the lysates of PC3 cells transfected with the indicated constructs. Cells were treated with MG132 (30 μM) for 6 h before collecting and lysis in denaturing buffer. h, Immunoblot analysis of WCL and immunoprecipitate derived from 293T cells transfected with indicated constructs and treated with MG132 (10 μM) for 12 h before collecting. i, Immunoblot of WCL derived from MDA-MB-231 PD-L1-knockout cells stably expressing wild-type PD-L1 or Δ283–290 and T290M PD-L1 mutants, with empty vector used as a negative control. j, Immunoblot analysis of WCL derived from HEK293T cells transfected with wild-type HA–PD-L1 or the T290M mutant. Cells which were treated with CHX for indicated times before collecting. k, Immunoblot of WCL and Ni–NTA pull-down products derived from the lysates of PC3 cells transfected with the indicated constructs. Cells were treated with MG132 (30 μM) for 6 h before collecting and lysis in denaturing buffer. l, Immunoblot of WCL derived from 293T cells transfected with indicated constructs.
Extended Data Figure 5 SPOP negatively regulates PD-L1 protein stability in a poly-ubiquitination-dependent manner.
a–c, Immunoblot analysis of WCL derived from 293T cells transfected with indicated constructs. d, e, Immunoblot analysis of WCL derived from 293T cells transfected with indicated constructs. Thirty-six hours after transfection, cells were treated with 20 μg ml−1 CHX for indicated length of time (d). The PD-L1 protein abundances were quantified using ImageJ (e). f, Immunoblot of WCL and Ni–NTA pull-down products derived from the lysates of PC3 cells transfected with the indicated constructs. Cells were treated with MG132 (30 μM) for 6 h before collecting and lysis in the denaturing buffer. g, A schematic illustration of SPOP with MATH and BTB domains interacting with substrate and cullin 3, respectively. h, Immunoblot analysis of WCL and immunoprecipitate derived from HEK293T cells transfected with indicated constructs and treated with MG132 (10 μM) for 12 h before collecting. i, Immunoblot analysis of WCL derived from HEK293T cells transfected with indicated constructs. j, qRT–PCR analysis of relative mRNA levels of PD-L1 (also known as Cd274) from Spop+/+ and Spop−/− MEFs. Data are mean ± s.d., n = 5. NS, not significant, two-tailed t-test. k, Immunoblot analysis of WCL derived from PC3 cells infected with indicated lentiviral shRNAs against SPOP and selected with puromycin (1 μg ml−1) for 72 h before collecting. l, m, Immunoblot analysis of WCL derived from C4-2 cells with depletion of SPOP using sgRNA, treated with CHX for indicated time points before collecting (l). The PD-L1 protein abundance was quantified using ImageJ (m). n, o, Immunoblot analysis of WCL derived from LNCaP cells stably expressing shAR or shERG as well as shScr as a negative control. p, q, Immunoblot analysis of WCL derived from DU145 cells stably expressing shTRIM24 or shDEK as well as shScr as a negative control. r–u, Immunoblot analysis of WCL derived from C4-2 wild-type SPOP and SPOP−/− cells that stably expressed shAR, shERG, shTRIM24, or shDEK as well as shScramble.
Extended Data Figure 6 Cancer-derived SPOP mutations fail to promote PD-L1 degradation.
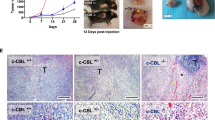
a, The mutation frequency (mutated cases/total cases) of SPOP across 24 cancer types from The Cancer Genome Atlas (TCGA) database. Mutations are categorized as occuring in the MATH domain, in the BTB domain or at any other position of the gene, including untranslated regions (UTRs). Because some patient cases contain mutations of two or three categories, the proportions of the colours are allocated mutation-wise, instead of case-wise. b, The distribution of mutation positions of SPOP in 24 cancer types from the TCGA database. Mutations without altering amino acid sequence have been discarded. c, Immunoblot analysis of WCL derived from HEK293T cells transfected with indicated constructs. d, The mutation frequency (mutated cases/total cases) of PD-L1 across 19 cancer types from the TCGA database. e, Oncoplot of PD-L1 and SPOP across all 39 cancer types in the TCGA database. Only PD-L1 or SPOP mutant tumours are shown; only mutations or truncations in the C-terminal tail of PD-L1 or in the MATH domain of SPOP are counted. f, Immunoblot of WCL derived from B16-F10 mouse tumour cell line stably expressing the indicated SPOP constructs. g, h, Growth curve and cell cycle profile of B16-F10 cells stably expressing wild-type SPOP and the F102C mutant, empty vector was used as a negative control. i, Cell cycle profile of 22Rv1 cells stably expressing wild-type SPOP and the F102C mutant, empty vector was used as a negative control. j, Relative cell-surface PD-L1 expression in 4T1-implanted tumours expressing wild-type SPOP or the F102C mutant measured using FACS analysis. n = 5 mice per experimental group. k, B16-F10 tumours derived from cells stably expressing wild-type SPOP or the F102C mutant and implanted in C57BL/6 mice were dissected and photographed after euthanization of mice. l, FACS analysis of the absolute number of CD3+ TILs from 4T1 tumours stably expressing wild-type SPOP or the F102C mutant. n = 5 mice per experimental group. m, B16-F10 tumour cells stably expressing wild-type SPOP or the F102C mutant were implanted into C57BL/6 mice. Tumour-bearing mice were treated with anti-PD-L1 antibody; tumours were then dissected and photographed. n = 7 mice per experimental group. n, The weight of B16-F10 cell derived tumours implanted in C57BL/6 mice that were subsequently treated with anti-PD-L1 antibody. 12 mice per experimental group. o, FACS analysis of relative cell surface expression of PD-L1 in implanted tumours derived from B16-F10 cells expressing wild-type SPOP or the F102C mutant. Animals were treated with anti-PD-L1 antibody. n = 5 mice per experimental group. p, FACS analysis of the absolute number of CD3+ TILs from B16-F10 tumours stably expressing wild-type SPOP or the F102C mutant in mice treated with anti-PD-L1 antibody. n = 7 mice per experimental group. q, B16-F10 cells stably expressing wild-type SPOP or the F102C mutant were inoculated into Tcra−/− mice. After two weeks, tumours were dissected and photographed. n = 7 mice per experimental group. r, Tumour weights of B16-F10 cells stably expressing wild-type SPOP or the F102C mutant and implanted into Tcra−/− mice were dissected and recorded. n = 7 mice per experimental group. s, FACS analysis of relative cell surface expression of PD-L1 from B16-F10 tumours stably expressing wild-type SPOP or the F102C mutant arising in Tcra−/− mice. n = 7 mice per experimental group. Data are mean ± s.d. *P < 0.05, **P < 0.01, ***P < 0.001, NS, not significant, two-tailed t-test.
Extended Data Figure 7 Validation of anti-PD-L1 and anti-CD8 antibodies through using PD-L1 knockout or shCD8 cells.
a, Immunoblot analysis of WCL derived from PD-L1 knockout MDA-MB-231 cells. b, Immunofluorescence of wild-type and PD-L1 knockout MDA-MB-231 cells using the anti-PD-L1 antibody. c, d, Immunochemistry using the anti-PD-L1 antibody of wild-type and PD-L1 knockout MDA-MB-231 cells from cultures on glass slides (c) or from implanted tumours (d). e, f, Immunoblot analysis using the anti-CD8 antibody of WCL derived from HBP-ALL (e) or KE37 (f) cells stably expressing shCD8 as well as shScr as a negative control. g, h, Immunohistochemistry using the anti-CD8 antibody for HBP-ALL (g) or KE37 (h) cell pellets stably expressing shCD8 or shScr as a negative control. Scale bars, 50 μm.
Extended Data Figure 8 Depletion of FZR1, but not CDC20, prolongs SPOP proteins stability, which is simultaneously coupled with a decrease in PD-L1 protein levels.
a–c, Immunoblot analysis of WCL derived from SPOP-knockout (a), CDC20-knockdown (b) or FZR1-knockdown (c) HeLa cells. d, Immunoblot analysis of WCL and immunoprecipitate derived from 293T cells transfected with indicated constructs and treated with MG132 (10 μM) for 12 h before collecting. e, Immunoblot analysis of WCL and immunoprecipitate derived from HeLa cells treated with MG132 (10 μM) for 12 h before collecting. f, A sequence comparison of D-box motifs (RXXLXXXXN) in SPOP derived from different species. g, Immunoblot analysis of WCL derived from HeLa cells transfected with indicated constructs. h, i, Immunoblot analysis of WCL derived from 293T cells transfected with indicated constructs. Thirty-six hours after transfection, cells were treated with CHX at indicated time points before collecting (h). The protein abundances of wild-type SPOP and ΔRXXL mutant were quantified using ImageJ (i).
Extended Data Figure 9 Cyclin D–CDK4-mediated phosphorylation of SPOP at the Ser6 residue promotes its binding with 14-3-3γ to reduce its poly-ubiquitination and subsequent degradation by APC–FZR1.
a, A sequence comparison of conserved SP sites and putative 14-3-3γ binding motif in SPOP. b, Immunoblot analysis of WCL and immunoprecipitate derived from 293T cells transfected with indicated constructs and treated with MG132 (10 μM) for 12 h before collecting. c, d, In vitro kinase assays with recombinant Rb and SPOP as substrates and cyclin D1–CDK4, cyclin D2–CDK4 and cyclin D3–CDK4 as kinase complexes were performed. BSA was used as a negative control where indicated. e, Immunoblot analysis of WCL and immunoprecipitate derived from MDA-MB-231 cells transfected with indicated constructs, with and without palbociclib treatment (1 μM) for 12 h. f, Streptavidin bead pull-down assay for biotin-labelled SPOP peptide with and without phosphorylation at the Ser6 residue to examine its in vitro association with 14-3-3γ. g, Immunoblot analysis of WCL and GST pull-down precipitates derived from 293T cells transfected with indicated constructs and treated with MG132 (10 μM) for 12 h before collecting. h, i, Immunoblot analysis of WCL and immunoprecipitates derived from 293T cells transfected with indicated constructs and treated with MG132 (10 μM) for 12 h before collecting. j, k, Immunoblot analysis of WCL derived from 293T cells transfected with indicated constructs. 36 h after transfection, cells were treated with 20 μg ml−1 CHX at indicated time points (j). The protein abundance of wild-type SPOP and the S6A mutant were quantified using ImageJ (k). l, p, Immunoblot of WCL and Ni–NTA pull-down products derived from the lysates of PC3 cells transfected with the indicated constructs. Cells were treated with MG132 (30 μM) for 6 h before collecting and lysis in denaturing buffer for subsequent assays. m–o, Immunoblot analysis of WCL and immunoprecipitates derived from 293T cells transfected with indicated constructs and treated with MG132 (10 μM) with or without palbociclib (1 μM) for 12 h before collecting. q–s, Immunoblot of WCLs derived from PC3 (q), BT549 (r) and HeLa (s) cells stably expressing sh14-3-3γ as well as shScr as a negative control. t, Immunoblot of WCL derived from HeLa cells stably expressing shScr or sh14-3-3γ synchronized in M phase by nocodazole treatment before release back into the cell cycle for the indicated times.
Extended Data Figure 10 Combination therapy of anti-PD-1 antibody and CDK4/6 inhibitor in MC38 or CT26 colon cancer mouse model.
a, A schematic of the treatment plan for mice bearing subcutaneous MC38 and CT26 tumours. Female C57BL/6 mice (for MC38) or BALB/c mice (for CT26) were implanted with 1 × 105 MC38 or CT26 cells subcutaneously and received one of four treatments: control antibody treatment, anti-PD-1 antibody treatment, CDK4/6 inhibitor (palbociclib) treatment, or anti-PD-1 antibody plus CDK4/6 inhibitor combination treatment. b, MC38 implanted tumour-bearing mice were enrolled in different treatment groups as indicated. Tumour volumes of mice treated with control antibody (n = 15), anti-PD-1 antibody (n = 15), palbociclib (n = 14) or anti-PD-1 antibody plus palbociclib combined therapy (n = 12) were measured every three days and plotted individually. Experiment repeated twice. c, Kaplan–Meier survival curves for each treatment group demonstrate the improved efficacy of combining PD-1 antibody with palbociclib. *P < 0.05, Gehan–Breslow–Wilcoxon test. Experiment repeated twice. d, e, g, i, The absolute numbers of CD3+, CD4+, CD8+, granzyme B+ or IFNγ+ TILs in implanted MC38 tumours from mice treated with indicated agents was analysed by FACS. Control: n = 8, palbociclib: n = 10, anti-PD-1 antibody: n = 9, palbociclib and anti-PD-1 antibody: n = 8. f, h, j, The percentage of CD4+, CD8+ in CD3+ TILs cells in implanted MC38 tumours from mice treated with indicated agents was analysed by FACS. Control: n = 8, palbociclib: n = 10, PD-1 antibody: n = 9, palbociclib and anti-PD-1 antibody: n = 8. k, A working model of how PD-L1 protein stability is regulated by the cyclin D–CDK4–SPOP–FZR1 signalling pathway. Cyclin D–CDK4 negatively regulates PD-L1 protein stability largely through phosphorylating its upstream physiological E3 ligase SPOP. This phosphorylation promotes SPOP binding to 14-3-3γ, which subsequently disrupts FZR1-mediated destruction of SPOP. Thus, CDK4/6 inhibitor treatment increases PD-L1 protein levels by inhibiting cyclin D–CDK4-mediated phosphorylation of SPOP to promote its degradation by APC/C. The rise in PD-L1 could present a severe clinical problem for patients receiving CDK4 inhibitor treatment and could be one of the underlying mechanisms accounting for CDK4 inhibitor resistance via evasion of immune surveillance checkpoints. Hence, our work provides a molecular mechanism as well as the rationale for the combination of PD-L1 blockade treatment and CDK4/6 inhibitors as a more efficient anti-cancer clinical option. Data are mean ± s.d. *P < 0.05, **P < 0.01, ***P < 0.001, two-tailed t-test.
Supplementary information
Supplementary Figure 1
This file contains the source data for gels in Figures 1-4 and Extended Data Figures 1-9. (PDF 4329 kb)
Rights and permissions
About this article

Cite this article
Zhang, J., Bu, X., Wang, H. et al. Cyclin D–CDK4 kinase destabilizes PD-L1 via cullin 3–SPOP to control cancer immune surveillance. Nature 553, 91–95 (2018). https://doi.org/10.1038/nature25015
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/nature25015
This article is cited by
-
ESCO2’s oncogenic role in human tumors: a pan-cancer analysis and experimental validation
BMC Cancer (2024)
-
Quantified pathway mutations associate epithelial-mesenchymal transition and immune escape with poor prognosis and immunotherapy resistance of head and neck squamous cell carcinoma
BMC Medical Genomics (2024)
-
CDKL1 potentiates the antitumor efficacy of radioimmunotherapy by binding to transcription factor YBX1 and blocking PD-L1 expression in lung cancer
Journal of Experimental & Clinical Cancer Research (2024)
-
Systematic investigation of chemo-immunotherapy synergism to shift anti-PD-1 resistance in cancer
Nature Communications (2024)
-
Unique vulnerability of RAC1-mutant melanoma to combined inhibition of CDK9 and immune checkpoints
Oncogene (2024)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
