
Abstract
Tumour neoantigens arising from cancer-specific mutations generate a molecular fingerprint that has a definite specificity for cancer. Although this fingerprint perfectly discriminates cancer from healthy somatic and germline cells, and is therefore therapeutically exploitable using immune checkpoint blockade, gut and extra-gut microbial species can independently produce epitopes that resemble tumour neoantigens as part of their natural gene expression programmes. Such tumour molecular mimicry is likely not only to influence the quality and strength of the body’s anti-cancer immune response, but could also explain why certain patients show favourable long-term responses to immune checkpoint blockade while others do not benefit at all from this treatment. This article outlines the requirement for tumour neoantigens in successful cancer immunotherapy and draws attention to the emerging role of microbiome-mediated tumour neoantigen mimicry in determining checkpoint immunotherapy outcome, with far-reaching implications for the future of cancer immunotherapy.
Similar content being viewed by others

Background
One of the hallmarks of cancer is the overly high proliferative capacity of tumour cells, which can be therapeutically exploited using classical cytotoxic chemotherapy. The merit of anti-proliferative drugs in the debulking of large tumours is beyond dispute, but these agents are molecularly unselective for cancer, and invariably also damage healthy tissue such as bone marrow and the colonic mucosa.1 Targeted drugs have successfully tackled this issue and facilitated cancer-predominant therapy with a superior safety profile (e.g. trastuzumab for HER2+ breast cancer,2 crizotinib for EML4-ALK+ lung cancer,3 imatinib for BCR-ABL+ leukaemia4). However, most targeted drugs are not exclusively specific for cancer, because they usually inhibit wild-type versions of proteins, which can often be expressed in a wide range of tissues (e.g. HER2 is also expressed in cardiomyocytes,5 and crizotinib6 and imatinib7 inhibit wild-type ALK and ABL tyrosine kinases, respectively, as well as the underlying cancer-specific gene fusions), and some small-molecule inhibitors, particularly first-generation agents, are multi-specific, thus showing additional on-target activity (e.g. crizotinib also inhibits MET and ROS1,8,9 and imatinib also inhibits c-kit and platelet-derived growth factor receptor10). Chimeric antigen receptor T cells also lack molecular specificity for cancer as their engineered affinities generally recognise wild-type antigens expressed on non-transformed cells as well as transformed cells, potentially leading to on-target, off-tumour toxicity (e.g. CD19 is expressed on both the malignant and healthy B-cell lineage).11,12
Despite the growing appreciation of epigenetic regulation13 and well-documented evidence for infectious contributions to tumour development (e.g. human papilloma virus14 and Helicobacter pylori15), the dogma that ‘cancer is a genetic disease’ still holds true.16 The stepwise accumulation of mutations not only facilitates and exacerbates malignancy, but also generates a molecular fingerprint—comprised of tumour neoantigens—with definite specificity for cancer. While some tumour neoantigens are recurrent among cancer entities (e.g. KRAS position 12 mutations), the overall molecular fingerprint of a cancer is unique in every patient irrespective of tumour type. This collective of cancer-specific molecular aberrations, commonly termed the ‘tumour neoantigenome’, is an ideal target for cell-mediated immunity and cancer immunotherapy, with arguably the highest possible specificity for cancer considering the natural selectivity of cytotoxic, adaptive immune cells.17,18,19 Exogenously derived antigens can also be regarded as ‘tumour neoantigens’20 but, compared with endogenous neoantigens, the presence of viral or bacterial antigens is limited to relatively few cancer entities (e.g. cervical cancer, gastric cancer, hepatocellular carcinoma, head and neck squamous cell carcinoma (HNSCC)).21 In addition, infection-related cancers still carry endogenous neoantigens,22,23,24 and some of these malignancies can be prevented with vaccines.25 This article will therefore prioritise endogenous tumour neoantigens arisen from mutations in host cells.
Although the presence of tumour neoantigens can discriminate cancer cells from ‘normal’ cells, epitopes produced by gut and extra-gut microbial species as part of their natural gene expression programmes can resemble tumour neoantigens—a phenomenon known as molecular mimicry. Molecular mimicry of tumour neoantigens by microbial species is likely to influence the host anti-cancer immune response through neoantigen-reactive T cells, and could explain why certain patients show favourable long-term responses to immune checkpoint blockade while others do not benefit at all from this treatment—tumour neoantigens are a mechanistic requirement for successful cancer immunotherapy.
In this article, we will introduce the concept of microbial tumour neoantigen mimicry emanating from the huge overall amount and diversity of microbial genetic sequences in the human body, and elaborate on the potential significance of such mimicry for checkpoint immunotherapy responsiveness and outcome.
Tumour neoantigens
Tumour neoantigens mostly constitute de novo mutations that arise in transformed cells when tumour DNA is altered through non-synonymous point mutations, gene translocations, or insertions and deletions (indels)20 and are excluded from central tolerance20—that is, they are not included in the mechanism used by the host to avoid attack by the immune system. Tumour neoantigens can also, in some cases, arise from the cancer-cell-specific usage of the epigenetic machinery, which can lead to the re-expression of silenced genes, the occurrence of unique splice or structural variants, and/or read-through transcription generating chimeric or intergenic products.20,26,27
Immunological visibility and immune evasion
Generally, immunological visibility of tumour neoantigens requires protein synthesis and a certain stability of the neoantigen product. Following the release of cytoplasmic content during cell death, for example, tumour neoantigens expressed as proteins will be taken up by tissue-patrolling antigen-presenting cells (APCs), which will then migrate to the tumour-draining lymph node and present the molecular tumour fingerprint to CD8+ naive T cells (cross-priming).28,29,30 In turn, interactions between T cells with a cognate T cell receptor and the neoantigen-positive APCs induce clonal expansion and effector/memory differentiation;31 cytotoxic tumour killing by neoantigen-specific T cells results in cancer cell lysis, which might generate an ‘in situ vaccination effect’ such that beneficial epitope spreading might occur.32 In an ideal scenario, sustained anti-cancer immunity will ensue protection from cancer re-growth.
However, clinical evidence indicates that such protective anti-cancer immunity is rare among cancer patients, owing to immune evasion caused by various cancer-cell-intrinsic33,34,35 and microenvironmental mechanisms.1,36 In terms of cancer-cell-intrinsic mechanisms, downregulation of the major histocompatibility complex class I (MHC I) presentation pathway, which is essential for immune recognition and CD8+-cytotoxic-cell-mediated killing, is often observed in tumour cells, as is the increased expression of immune checkpoint molecules, which function as a ‘brake’ on the immune system, controlling the duration and extent of immune responses, maintaining self-tolerance and preventing autoimmunity.
Immune checkpoint inhibitors
Immune checkpoint inhibitors (ICIs) that target checkpoint components, such as programmed death receptor (PD-1) and its ligand programmed cell death ligand 1 (PD-L1), or cytotoxic T-lymphocyte-associated protein 4 (CTLA-4), or other inhibitory molecules can partly release this immune system brake and have shown impressive long-term outcomes in many cancer patients.37,38,39,40 Mechanistically, ICIs neutralise negative feedback from immunological checkpoints or increase immune-stimulatory signalling, thus reinvigorating the endogenous tumour-specific T cell response. For CTLA-4-targeted immunotherapy, it has been shown that the clinical mode of action is likely to involve therapy-enhanced T cell priming, which broadens the tumour-specific CD8+ T cell response.41 Data from anti-CTLA-4-treated melanoma patients demonstrated a significant increase in the number of newly detected tumour-specific CD8+ T cell responses after treatment, whereas pre-existing virus- and tumour-specific T cell responses remained unchanged.41 Similar findings were reported for ICIs that target PD-1, thus altogether suggesting that pre-existing tumour-reactive T cells have a limited reinvigoration potential, which is outcompeted by de novo arising T cell specificities (clonal replacement).42 The poor reinvigoration capacity of pre-existing tumour-specific T cells might indicate a partially exhausted cell state maintained independently from checkpoint signalling, which potentially limits the use of ICIs in certain patients.
The clinical significance of ICIs is tremendous, as these agents have shown unprecedented responses even in advanced-stage tumours,43 and have established themselves as frontline therapeutics in a broad variety of solid tumour types.44,45 However, the speed of further development of ICI-based immunotherapy has started to decelerate, and significant challenges lie ahead, including the need for adapted clinical trial design and customisation of clinical safety and efficacy endpoints to more specifically account for the characteristic features of ICIs compared with classical anti-cancer drugs.46
The relevance of tumour neoantigens in medical oncology
The relevance of tumour neoantigens in medical oncology is highlighted by the positive association between tumour mutational burden (TMB) and the response to ICI treatment.47,48,49,50,51,52,53 Accordingly, tumour entities with a typically high TMB (e.g. lung cancer, skin cancer, microsatellite-instable cancers) respond significantly better to ICI-based immunotherapy than do cancers with a comparatively low TMB (e.g. breast cancer, prostate cancer).54 Moreover, immunological treatments that target the tumour neoantigenome through reinvigoration (in the case of ICIs) or induction (for vaccines) of antigen-specific CD8+ T cells do not rely solely on the ‘driver’ nature of mutations, as passenger mutations can be equally immunogenic and thereby also serve as targets for immune destruction;19,55,56 by contrast, targeted treatment options require that the respective target molecule is causal for tumour growth and/or progression (‘oncogene addiction’).57,58
Thus, tumour neoantigens arising from cancer-specific mutations are attractive targets for cancer immunotherapy, which has shown proven clinical relevance and important conceptual advantages over other types of cancer treatment that do not exploit the natural adaptability and versatility of the immune system.
Checkpoint blockade responsiveness
Although ICIs have revolutionised the treatment of various solid tumour types and enabled unprecedented long-term remissions, a significant number of patients do not benefit from these therapies, owing to primary, adaptive, or acquired resistance mechanisms59,60 shown by tumours. Primary resistance mechanisms include the absence of a relevant number of immunogenic tumour antigens, defects in the antigen processing and presenting machinery, and insufficient T cell infiltration.59 Adaptive resistance to cancer immunotherapy refers to mechanisms by which cancer cells adapt to attacking immune cells, for instance through downregulation of MHC class I or upregulation of PD-L1.59 Acquired resistance refers to a clinical scenario where a patient initially responds to immunotherapy but later on progresses or relapses; oftentimes, this will involve Darwinian selection of resistant clones present already before treatment start.59
Only a small proportion of patients with non-small cell lung cancer (NSCLC), for example, respond favourably to ICI treatment.61,62,63 The time lost in treating poorly responding or refractory patients, along with the potential toxicity64,65 and high associated costs of ICI treatment,66,67 demands that better tools and predictive markers are established to discriminate patients who are likely to respond to treatment from those who are not likely to respond. More importantly, identifying the underlying mechanisms of non-responsiveness will be instrumental for the development of therapies that work in combination with ICIs to resensitise tumours to checkpoint inhibition.
Current markers for predicting responsiveness
Aside from TMB as a surrogate indicator for tumour neoantigens, the extent of tumour T cell infiltration at baseline and the expression of the respective ICI target (e.g. PD-1/PD-L1) at the protein level represent the most established markers for predicting the responsiveness to checkpoint blockade.50,68,69 However, none of these markers is perfect and they are currently unable to predict treatment response with clinically sufficient precision.70,71 As an example, the predictive power of baseline tumour T cell infiltration might be reduced or lost if these T cells are exhausted or otherwise non-functional. Similarly, a high TMB indicating increased statistical odds for the presence of tumour neoantigens does not predict whether these cancer-specific antigens are immunogenic to an extent that is therapeutically relevant. Finally, the proven presence of the ICI target protein only indicates that the substrate is present to inhibit, but not whether, or to what extent, this substrate is causal for tumour growth. Therefore, novel and more reliable predictive markers are desirable to refine patient stratification and further optimise ICI treatment.
The significance of the gut microbiome in the responsiveness to ICI
Within the past decade, evidence has suggested an important role for the gut microbiome in determining the responsiveness to checkpoint immunotherapy. Two articles published in 2015 demonstrated that commensal Bifidobacterium72 and distinct Bacteroides species73 can promote anti-cancer immunity during ICI treatment. Accordingly, co-treatment with antibiotics reduces the clinical activity of checkpoint immunotherapy,74,75,76,77,78 whereas faecal microbiome transplantation (FMT) from ICI-responding patients to germ-free or antibiotic-treated mice partly rescues the anti-cancer effects of ICI treatment, dependent on the presence of A. muciniphila Gram-negative bacteria.79 Although these data can offer possibilities for straightforward therapeutic development (e.g. oral supplementation with relevant bacterial strains, FMT from responding patients), the mechanisms responsible for these observations remain elusive and a deeper understanding of the underlying biology is desirable.80
Immunological mimicry of tumour neoantigens by microbial peptide products in ICI responsiveness
A likely explanation for the functional significance of the gut microbiome in checkpoint immunotherapy responsiveness is immunological mimicry of tumour neoantigens by microbial peptide products81,82,83,84,85 (Fig. 1). Support for this hypothesis comes from the observation that the efficacy of anti-CTLA-4 treatment depends on antigen-specific immune reactivity against certain bacterial strains, including B. fragilis. Accordingly, if B. fragilis is experimentally lacking (in germ-free or antibiotic-treated mice), anti-CTLA-4 treatment efficacy can be restored by oral gavage of B. fragilis, immunisation with B. fragilis-derived products or adoptive transfer of T cells specific for B. fragilis.73 It is worth mentioning in this context that a 2020 publication on cancer immunotherapy targeting CD47 (another checkpoint molecule) found that the accumulation of gastrointestinal bacteria in subcutaneously grown tumours was critical for the treatment response.86 In addition, another study showed that intratumoural accumulation of bacteria is common among cancer entities, with bacterial compositions being mostly biased towards intracellular bacteria that are capable of triggering CD8+ T cell responses.87 These results highlight the far-reaching effects of gastrointestinal microbes across organ systems/compartments and clearly emphasise the significance of the local tumour microbiome for responsiveness to cancer immunotherapy. Aside from the well-known gastrointestinal microbiome, a rich flora of microbial communities can also be found in the respiratory tract88,89 as well as on other various mucosal surfaces and non-mucosal surfaces of the human body.90,91,92 Thus, certain carcinomas (e.g. NSCLC, HNSCC, cervical cancer) are likely to be influenced by a distinct tumour-associated habitat that might also shape the response to ICI therapy.

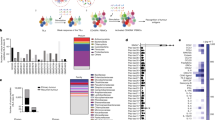
Depicted are important clinical parameters known to influence the response to checkpoint-based immunotherapy. The expression of the targets of immune checkpoint inhibitors (ICIs), the level of tumour mutational/neoantigen burden and the extent of T cell infiltration at baseline are established clinical predictors of ICI treatment response; the molecular contribution of the (gut) microbiome, however, is less well-studied. Epitope mimicry of tumour neoantigens is likely to explain the functional importance of microbial species in checkpoint immunotherapy responsiveness and arguably deserves prioritisation for scientific elaboration efforts. Note the interdependence of the various checkpoint-predictive markers and parameters, indicated with dashed lines. In brief, cancer cell-specific mutations are a requirement for tumour neoantigens, the number of which is thought to correlate with the overall tumour mutational burden. Both tumour neoantigens and tumour mutational burden influence the infiltration of tumours by T cells and are further associated with checkpoint molecule expression. The host microbiome also modulates the response to checkpoint blockade and is connected with tumour neoantigens through molecular mimicry. PD-L1, programmed cell death ligand 1.
Convincing evidence therefore suggests that the outcome of ICI-based cancer immunotherapy is influenced, at least partially, by the composition and functional quality of the (distant) gut- and the (local) tumour microbiome.
Tumour neoantigens and tumour neoantigen mimicry
Cancer immune surveillance is, to a large extent, carried out by CD8+ T cells. Molecularly, these cells recognise tumour neoantigens typically as 8–11-mer peptides93,94 bound to MHC class I on the tumour cell surface. Random 11-mer permutation of the 20 proteinogenic amino acids in humans (not counting atypical amino acids such as selenocysteine) can generate more than 1014 theoretical combinations, although the number of possible tumour neoantigens is correspondingly smaller considering that many of these sequence combinations will already be part of the host genome.
How does microbial mimicry to tumour antigens arise?
Many microbes that colonise the human body can synthesise all 20 proteinogenic amino acids of humans,95,96 and viruses can harness the host cell translational machinery to exclusively use the endogenous amino acid repertoire. It is believed that the human gastrointestinal tract harbours approximately 1014 microbes, which, together, equate to roughly 1000 times the number of cells and 10000 times the DNA content of the human body.97 Exact mathematical modelling is beyond the scope of this article, but it is with definite statistical likelihood that numerous individual tumour neoantigens are immunologically mimicked by peptide products of gut and extra-gut microbial species: identical peptide mimics comprise the same amino acid sequence as the corresponding tumour neoantigens and result in identical T cell epitopes, whereas ‘notably similar’ peptide mimics contain the same epitope core but differ in their flanking region or length, thus altering antigen presentation efficiency and/or generating T cell epitopes with nuance-like specificity alterations.98,99 Importantly, the actual likelihood of mimicry might be smaller than theoretically possible considering that microbial genomes encode functional proteins whose sequences might not be randomly structured which might lower the number of actually possible sequence permutations. In addition, not every neoantigen-mimicking microbial peptide will reach a concentration that is high enough for immunological recognition and modulation of the anti-cancer immune response.
The role(s) of tumour neoantigen microbial mimicry
The abundance of class-switched IgA antibodies with high affinities for microbial species in the gut microenvironment provides clear evidence for APC-dependent, humoral responses elicited and matured in gut-associated lymphoid tissue and/or regional lymph nodes.100,101 Such microbial-antigen-containing APCs can also cross-present their phagocytosed cargo via MHC class I to induce cellular immune responses.102,103 Furthermore, intracellular microbes present in both cancer and immune cells87 can trigger direct CD8+ T cell responses in the tumour microenvironment (TME). Tumour neoantigen mimicry by microbial species is therefore likely to produce T cell clones of both the CD8 and CD4 lineage that are tumour-reactive despite never having previously seen actual tumour antigens. Such independent emergence of T cell populations with overlapping specificity can lead to functional immunological crosstalk that shapes overall anti-cancer immunity and influences the outcome of checkpoint-based immunotherapy.81,82,83,84
Currently, the net effect of such immune interaction is unclear. Generally, the gastrointestinal tract and other body surfaces are considered a rather tolerogenic environment, an idea that is consistent with the notion of productive interplay with commensals under homoeostatic conditions104,105 as well as the default unresponsiveness to food proteins in healthy individuals (oral tolerance).106,107 However, it is unknown how compartment-specific microbial tolerance influences neoantigen-directed tumour immune surveillance (even more so during perturbations such as dysbiosis or infection). Although immune tolerance to commensals and their tumour neoantigen mimics might intuitively be more associated with reduced anti-cancer immunity, evidence has shown that T cells that are specific for certain gut commensals serve immune-stimulatory functions during checkpoint-based immunotherapy.73 These data suggest that the ‘tolerogenic’ environment of the gut, including tolerance to neoantigen-mimicking commensals, can nevertheless support anti-cancer immunity based on yet-to-be-determined mechanisms; conceivable means include compartment-specific effects as well as temporary disruptions of epithelial barrier function leading to translocation into the systemic circulation and subsequent failure of tolerance.
In further support of clinically relevant tumour neoantigen mimicry, long-term survivors of pancreatic cancer show tumour-neoantigen-specific T cell clones with predicted cross-reactivity to microbial epitopes.81 In these patients, immunogenic neoantigens can be identified using a quality fitness model that takes into account differential presentation and homology to microbial-derived peptides.81 Of note, ‘high-quality’ neoantigens are selectively lost upon metastatic disease progression, suggesting immunoediting and, ultimately, immunological escape of clinically relevant tumour neoantigens.81
Immune-modulatory activity of microbial metabolites
The microbiome-derived metabolome refers to the aggregate of individual microbial metabolites in a given compartment, several of which, including short-chain fatty acids108,109 and inosine,110 are known to have immune-modulatory activity and to influence cancer immunotherapy. Short-chain fatty acids are differentially expressed among immunotherapy responders and non-responders, and a high faecal abundance of acetic, propionic, butyric and valeric acid correlates with prolonged progression-free survival.108,109 The underlying mechanisms are yet to be fully elucidated, but are likely to involve altered T cell homoeostasis and differentiation as well as modulation of APC functions108,109 brought about by these short-chain fatty acids. Inosine as produced by Bifidobacterium spp. in the gut facilitates enhanced ICI treatment efficacy through a mechanism that involves the increased systemic translocation (owing to ICI-induced decreased gut barrier function) of inosine and subsequent activation of anti-cancer T cells.110 Considering the basic principles of immune activation and differentiation, it is very much conceivable that T cells, as well as their correspondingly induced cytokines, that recognise mimicked tumour neoantigens are subject to regulation by microbial metabolites.
Dissecting the role of microbial tumour neoantigen mimicry
At this stage, no conclusions on the role of microbial tumour neoantigen mimicry can be drawn; mechanistic studies in mice as well as consolidation in the clinical setting are warranted. Below we provide insight into how to experimentally address microbial tumour neoantigen mimicry to better understand responsiveness to checkpoint immunotherapy and optimise patient stratification. Molecular and immunological tools are available to define the contribution of gut microbes to the responsiveness of checkpoint immunotherapy and to systematically assess whether microbial tumour neoantigen mimicry acts to foster or impede protective anti-cancer immunity.
Generating mouse models of immunological tumour mimicry
Modern genetic engineering facilitates the expression of almost any gene of interest in prokaryotic and eukaryotic cells and is key to mechanistically dissecting the function of genes. Using this reverse genetics approach to introduce tumour neoantigens into gut commensals, mouse models of immunological tumour mimicry can be experimentally generated. To achieve this, the genetic sequences encoding tumour neoantigens need to be cloned into suitable commensals (e.g. Bifidobacterium, Bacteroides and Escherichia spp.) before the genetically modified bacteria are introduced into the gastrointestinal tract of immunocompetent mice using oral gavage; sustained bacterial engraftment might involve antibiotic conditioning.111 Tumour neoantigen mimicry can be enforced by transplanting tumour cells into bacteria-reconstituted mice, and the impact of such mimicry on anti-cancer immunity and immunotherapy responsiveness can be investigated (Fig. 2a). Syngeneic, orthotopic tumour models are desirable, as they most closely resemble the physiological setting especially in terms of immune compatibility and an authentic TME.112,113 A future option might involve the use of patient-derived, orthotopic xenograft models114 in conjunction with human haematopoiesis-reconstituted mice (e.g. the MISTRG strain115), provided that limitations with adaptive immune responses can be overcome in humanised mice.116

a Tumour cells of known neoantigen status and corresponding engineering of gut commensals can be used to generate mouse models of enforced microbial tumour neoantigen mimicry. The effects of such mimicry on checkpoint immunotherapy responsiveness and tumour growth can be investigated and the quality and strength of the anti-cancer immune response can be assessed. System perturbations such as co-treatment with antibiotics can be introduced to fine-tune the degree of tumour neoantigen mimicry in these well-defined models. b Using whole-exome sequencing and RNA sequencing of tumour and blood samples from cancer patients treated with immune checkpoint inhibitors, tumour neoantigens binding to respective MHC class I molecules can be identified using bioinformatics. In parallel, shotgun sequencing of stool samples can characterise the microbial metagenome and identify the antigenic sequences present in the gastrointestinal tract. Using this information, the degree of immunological similarity between tumour cells and gut microbes can be determined and summarised as ‘tumour antigenic similarity’ (TAS). Using TAS-based patient stratification, the impact of microbial tumour neoantigen mimicry on treatment response and survival can be evaluated. Companion immune profiling using methods such as enzyme-linked immunospot assay and T cell receptor sequencing can be used to corroborate clinical survival data and indicate the potential underlying mechanisms of treatment response or failure. ELISPOT enzyme-linked immunospot assay, NGS next-generation sequencing, TAS tumour antigenic similarity, TCR T cell receptor, WES whole-exome sequencing.
The mimicked (neo-)antigens can include surrogate tumour antigens such as minigene-encoded epitopes of viral glycoproteins117,118,119 and xenogeneic proteins,120,121,122 or bona fide tumour neoantigens that arise naturally during tumour evolution. Although surrogate tumour antigen models can provide unique opportunities to track antigen-specific immune responses (which is important for initial proof-of-concept studies),123,124 they are largely artificial and can have limited predictive and translational value.125,126 Conversely, tumour neoantigens from mutagen-induced (e.g. dimethylbenzathracene, azoxymethane, N-ethyl-N-nitrosourea, UV light127) or genetically-engineered tumours (e.g. KrasLSL-G12D; p53frt/frt lung adenocarcinoma,128,129 MMTV-PyMT130,131 or MMTV-neu132 mammary carcinoma, KrasLSL-G12D; p16lox/lox; Pdx1-Cre pancreatic ductal adenocarcinoma129,133) might be more relevant because they have evolved ‘naturally’ and gone through immune selection and immune-editing processes.134,135 Such engineered/autochthonous models will produce genetically heterogeneous tumours with a comparatively high frequency of protein-altering mutations and copy number variations.136 From such models, clonal tumour cell lines can be established, and the tumour neoantigens can be identified using next-generation sequencing at the DNA level and compared with autologous reference tissue/blood.18 RNA sequencing can determine the fraction of neoantigens putatively expressed, and peptide–MHC binding algorithms such as NetMHCpan137 can predict neoantigen immunogenicity.
Assessing the effects of immunological tumour neoantigen mimicry on immune response and tumour growth
To investigate the functional significance of immunological tumour neoantigen mimicry for ICI responsiveness and outcome, readouts addressing the quality and strength of the anti-cancer immune response as well as changes in tumour growth are required. Flow cytometry/fluorescence-activated cell sorting can help to delineate general immune (activation) patterns in tumour tissue, blood and lymph nodes, and can further facilitate neoantigen-specific T cell tracking and purification, making use of peptide–MHC multimer technology.119,123,138,139,140 The enzyme-linked immunospot assay can probe the neoantigen-specific reactivity of ex vivo stimulated T cell populations141,142 and T cell receptor sequencing can characterise the clonality and overall repertoire of intratumoural T cell subsets.143
Complementing these important immune-targeted readouts, the effects on functional tumour growth need to be assessed, and a number of scenarios should be explored: first, ‘continuous’ tumour growth resulting from a high dose of transplanted tumour cells; second, tumour engraftment emanating from a critically low dose of transplanted tumour cells;139,144 and, third, ‘metastatic’ growth in a relevant compartment.119 These scenarios will allow various clinical settings, including those of high tumour burden, minimal residual disease and oligometastatic disease, respectively, to be experimentally explored. System perturbations such as co-treatment with antibiotics to reduce or abrogate microbial tumour neoantigen mimicry could be envisaged for control purposes; here, the particular timing of antibiotic treatment should be carefully selected considering treatment duration as well as lagging phase and washout effects.75
Moving into humans…
Scrutinising and validating these preclinical results in humans are crucial for eventual clinical translation. To this end, tumour neoantigens need to be identified from tumour biopsy samples using the strategy outlined above,18 and the gastrointestinal microbiome composition needs to be characterised using metagenomic shotgun sequencing of stool samples.145 Dedicated bioinformatics interrogation can uncover the overlapping tumour neoantigen and microbial sequences, and provide an estimate for the overall ‘tumour antigenic similarity’. Correlating the degree of this similarity with clinical endpoints such as survival, treatment response and immune-related adverse events, a potential proxy for treatment efficacy,65 will demonstrate how microbial tumour neoantigen mimicry affects patient outcomes under ICI therapy (Fig. 2b). Complementary immune profiling using neoantigen-specific (e.g. peptide-MHC multimers, enzyme-linked immunospot assay) and -unspecific assays (e.g. flow cytometry, expression profiling, T cell receptor sequencing) will be important to identify the potentially underlying mechanisms.
Conclusion and perspectives
Improved patient stratification and resensitisation to ICIs with appropriate drugs or interventions is key to fulfilling the potential of these immunotherapeutic agents in medical oncology. Although ICI target expression and a high TMB mechanistically facilitate and predict checkpoint immunotherapy responsiveness, they have no further therapeutic (e.g. treatment-resensitising) potential. By contrast, the composition of the gut microbiome is more dynamically regulated146,147 and might be an attractive target for therapeutic manipulation, given its causal role in ICI responsiveness and treatment outcome.72,73,74,75,79
Several mechanisms, including discrepancies in the levels of some metabolites,148,149 might conceivably be responsible for modulating both natural81 and ICI-induced73,82,83,84 anti-cancer immunity; however, evidence suggests that molecular mimicry of tumour neoantigens by microbial species is likely to be an important factor. The net effect of tumour neoantigen mimicry on ICI treatment efficacy currently remains unknown and future mechanistic and clinical studies are clearly warranted. Nonetheless, the results of various studies that have investigated microbial composition in the context of checkpoint immunotherapy responsiveness support a model in which mimicry has favourable effects on anti-cancer immunity and ICI-related treatment outcome (Fig. 3). First, co-treatment with antibiotics (which reduces potential mimicry) and organism-wide germ elimination (which precludes mimicry) are well-known to curtail the efficacy of checkpoint-based immunotherapy73,74,75,79,150—with the limitation that clinical data are derived almost exclusively retrospectively, as prospective testing of an ICI–antibiotic combination is not really feasible due to ethical concerns.77 Second, a higher microbial diversity index (which increases the statistical odds for mimicry) is associated with immune activation and long-term survival of pancreatic cancer patients151 as well as a memory T cell signature and favourable response to PD-1-targeted immunotherapy in patients with NSCLC.152 Along these lines, the beneficial effects of therapeutic FMT from ICI-responding patients79 might be at least partially attributable to an increase in the overall microbial diversity, similar to the results seen for FMT in patients with inflammatory bowel disease.153,154

a Scenarios of reduced or absent microbial diversity (representing little/no mimicry) are associated with poor responsiveness to immune checkpoint inhibitors (ICIs), whereas scenarios of reinstated or higher microbial diversity (representing relevant/substantial mimicry) are associated with favourable ICI responsiveness. b Schematic showing the putative role of microbial tumour neoantigen mimicry in checkpoint-based immunotherapy. Microbial species mimicking tumour neoantigens can independently elicit tumour-reactive T cells based on antigen-presenting cell (APC) cross-priming or default APC pathways (extracellular microbes), or native ‘self’ antigen processing in infected host cells (intracellular microbes). As shown in the green box at the right-hand-side of the panel, neoantigen molecular mimicry can result in overlapping CD8+ (and CD4+) T cell specificities which might potentiate ICI therapy and contribute to protective cancer immune surveillance through reinvigoration of tumour neoantigen-specific immunity. ABx antibiotic treatment, APC antigen-presenting cell, FMT faecal microbiome transplantation, ICI immune checkpoint inhibitor.
Assuming a protective role for microbial tumour neoantigen mimicry, interventions aimed at supplementing probiotics and/or increasing microbial diversity represent rational therapeutic approaches that warrant clinical investigation.85 Importantly, the gut microbiome engages in crosstalk with the tumour microbiome such that therapeutic FMT will automatically also modulate the tumour microbial composition.151 Clinical research will show whether such interventions are sufficient to reinstate relevant tumour neoantigen mimicry or whether targeted engineering of commensals based on the tumour neoantigen landscape would be required. Such personalised, molecularly informed approaches are technically feasible, and could be achieved within a clinically reasonable time frame (approximately 6–8 weeks), provided that trial data can overcome safety concerns regarding the use of genetically-modified live bacteria.
Another promising anti-cancer strategy is the use of ICIs together with tumour-neoantigen-specific vaccines.18,141 Given the clarification of the definite role of tumour neoantigen mimicry in ICI responsiveness, this knowledge can help to complement MHC class I binding prediction137 and assist in the consideration of which neoantigens to choose for vaccination. In this way, the anti-cancer immune response could be (re-)directed specifically to molecularly mimicked neoantigens, provided that mimicry indeed shows beneficial effects on ICI treatment efficacy.
Immunogenic cancer cells adapt to the selective pressure of the immune system, conceptually traversing through three successive phases—elimination, equilibrium and escape135,155—by the process of immunoediting. Immunoediting of tumour neoantigens has been reported, and is associated with metastatic progression in pancreatic cancer.81 Other prominent mechanisms of cancer immune escape include somatic loss of HLA class I molecules (loss of heterozygosity (LOH))156 and defects in the MHC class I antigen processing and presenting machinery.157 Importantly, HLA class I LOH predicts poor overall survival in a subgroup of ICI-treated NSCLC patients and can be used as a complementary marker for checkpoint immunotherapy responsiveness to refine TMB-based stratification.158 In the context of tumour neoantigen mimicry and associated therapeutic interventions, HLA class I LOH needs to be considered as a potential confounder for tumour-microbiome cross-reactive T cell responses. While HLA class I LOH occurs spontaneously and is not preventable per se, any neoantigen-directed therapeutic intervention should ensure coverage of several HLA class I alleles to avoid dependence on a single genetic locus.
This review article has highlighted the crucial importance of tumour neoantigens for cancer immunotherapy and provided impetus for investigating the significance of microbial tumour neoantigen mimicry for checkpoint immunotherapy responsiveness and outcome. Immunologically informed considerations on ICI therapeutic use will optimise their clinical performance and set the stage for microbiome-targeted interventions to boost response rates and prolong survival.
References
Boesch, M., Baty, F., Rumpold, H., Sopper, S., Wolf, D. & Brutsche, M. H. Fibroblasts in cancer: defining target structures for therapeutic intervention. Biochim. Biophys. Acta Rev. Cancer 1872, 111–121 (2019).
Robert, N., Leyland-Jones, B., Asmar, L., Belt, R., Ilegbodu, D., Loesch, D. et al. Randomized phase III study of trastuzumab, paclitaxel, and carboplatin compared with trastuzumab and paclitaxel in women with HER-2-overexpressing metastatic breast cancer. J. Clin. Oncol. 24, 2786–2792 (2006).
Shaw, A. T., Kim, D. W., Nakagawa, K., Seto, T., Crino, L., Ahn, M. J. et al. Crizotinib versus chemotherapy in advanced ALK-positive lung cancer. N. Engl. J. Med. 368, 2385–2394 (2013).
O’Brien, S. G., Guilhot, F., Larson, R. A., Gathmann, I., Baccarani, M., Cervantes, F. et al. Imatinib compared with interferon and low-dose cytarabine for newly diagnosed chronic-phase chronic myeloid leukemia. N. Engl. J. Med. 348, 994–1004 (2003).
Gordon, L. I., Burke, M. A., Singh, A. T., Prachand, S., Lieberman, E. D., Sun, L. et al. Blockade of the erbB2 receptor induces cardiomyocyte death through mitochondrial and reactive oxygen species-dependent pathways. J. Biol. Chem. 284, 2080–2087 (2009).
Shaw, A. T., Yasothan, U. & Kirkpatrick, P. Crizotinib. Nat. Rev. Drug Discov. 10, 897–898 (2011).
Capdeville, R., Buchdunger, E., Zimmermann, J. & Matter, A. Glivec (STI571, imatinib), a rationally developed, targeted anticancer drug. Nat. Rev. Drug Discov. 1, 493–502 (2002).
Rothschild, S. I. & Gautschi, O. Crizotinib in the treatment of non–small-cell lung cancer. Clin. Lung Cancer 14, 473–480 (2013).
Sgambato, A., Casaluce, F., Maione, P. & Gridelli, C. Targeted therapies in non-small cell lung cancer: a focus on ALK/ROS1 tyrosine kinase inhibitors. Expert Rev. Anticancer Ther. 18, 71–80 (2018).
Savage, D. G. & Antman, K. H. Imatinib mesylate–a new oral targeted therapy. N. Engl. J. Med. 346, 683–693 (2002).
Rafiq, S., Hackett, C. S. & Brentjens, R. J. Engineering strategies to overcome the current roadblocks in CAR T cell therapy. Nat. Rev. Clin. Oncol. 17, 147–167 (2020).
Guedan, S., Ruella, M. & June, C. H. Emerging cellular therapies for cancer. Annu. Rev. Immunol. 37, 145–171 (2019).
Topper, M. J., Vaz, M., Marrone, K. A., Brahmer, J. R. & Baylin, S. B. The emerging role of epigenetic therapeutics in immuno-oncology. Nat. Rev. Clin. Oncol. 17, 75–90 (2020).
zur Hausen, H. Papillomavirus infections–a major cause of human cancers. Biochim. Biophys. Acta 1288, F55–F78 (1996).
Parsonnet, J., Friedman, G. D., Vandersteen, D. P., Chang, Y., Vogelman, J. H., Orentreich, N. et al. Helicobacter pylori infection and the risk of gastric carcinoma. N. Engl. J. Med. 325, 1127–1131 (1991).
Vogelstein, B., Papadopoulos, N., Velculescu, V. E., Zhou, S., Diaz, L. A. Jr. & Kinzler, K. W. Cancer genome landscapes. Science 339, 1546–1558 (2013).
Yarchoan, M., Johnson, B. A. 3rd, Lutz, E. R., Laheru, D. A. & Jaffee, E. M. Targeting neoantigens to augment antitumour immunity. Nat. Rev. Cancer 17, 209–222 (2017).
Hu, Z., Ott, P. A. & Wu, C. J. Towards personalized, tumour-specific, therapeutic vaccines for cancer. Nat. Rev. Immunol. 18, 168–182 (2018).
Gubin, M. M., Zhang, X., Schuster, H., Caron, E., Ward, J. P., Noguchi, T. et al. Checkpoint blockade cancer immunotherapy targets tumour-specific mutant antigens. Nature 515, 577–581 (2014).
Schumacher, T. N., Scheper, W. & Kvistborg, P. Cancer neoantigens. Annu. Rev. Immunol. 37, 173–200 (2019).
De Flora, S. & Bonanni, P. The prevention of infection-associated cancers. Carcinogenesis. 32, 787–795 (2011).
Qin, Y., Ekmekcioglu, S., Forget, M. A., Szekvolgyi, L., Hwu, P., Grimm, E. A. et al. Cervical cancer neoantigen landscape and immune activity is associated with human papillomavirus master regulators. Front. Immunol. 8, 689 (2017).
Castle, J. C., Uduman, M., Pabla, S., Stein, R. B. & Buell, J. S. Mutation-derived neoantigens for cancer immunotherapy. Front. Immunol. 10, 1856 (2019).
Leemans, C. R., Snijders, P. J. F. & Brakenhoff, R. H. The molecular landscape of head and neck cancer. Nat. Rev. Cancer 18, 269–282 (2018).
Saslow, D., Castle, P. E., Cox, J. T., Davey, D. D., Einstein, M. H., Ferris, D. G. et al. American Cancer Society Guideline for human papillomavirus (HPV) vaccine use to prevent cervical cancer and its precursors. CA Cancer J. Clin. 57, 7–28 (2007).
Smith, C. C., Selitsky, S. R., Chai, S., Armistead, P. M., Vincent, B. G. & Serody, J. S. Alternative tumour-specific antigens. Nat. Rev. Cancer 19, 465–478 (2019).
Varley, K. E., Gertz, J., Roberts, B. S., Davis, N. S., Bowling, K. M., Kirby, M. K. et al. Recurrent read-through fusion transcripts in breast cancer. Breast Cancer Res. Treat. 146, 287–297 (2014).
Roberts, E. W., Broz, M. L., Binnewies, M., Headley, M. B., Nelson, A. E., Wolf, D. M. et al. Critical role for CD103(+)/CD141(+) dendritic cells bearing CCR7 for tumor antigen trafficking and priming of T cell immunity in melanoma. Cancer Cell 30, 324–336 (2016).
Sanchez-Paulete, A. R., Teijeira, A., Cueto, F. J., Garasa, S., Perez-Gracia, J. L., Sanchez-Arraez, A. et al. Antigen cross-presentation and T-cell cross-priming in cancer immunology and immunotherapy. Ann. Oncol. 28, xii74 (2017).
Kurts, C., Robinson, B. W. & Knolle, P. A. Cross-priming in health and disease. Nat. Rev. Immunol. 10, 403–414 (2010).
Bousso, P. T-cell activation by dendritic cells in the lymph node: lessons from the movies. Nat. Rev. Immunol. 8, 675–684 (2008).
Brossart, P. The role of antigen-spreading in the efficacy of immunotherapies. Clin. Cancer Res. https://doi.org/10.1158/1078-0432.CCR-20-0305 (2020).
Cha, J. H., Chan, L. C., Li, C. W., Hsu, J. L. & Hung, M. C. Mechanisms controlling PD-L1 expression in cancer. Mol. Cell 76, 359–370 (2019).
Garrido, F., Aptsiauri, N., Doorduijn, E. M., Garcia Lora, A. M. & van Hall, T. The urgent need to recover MHC class I in cancers for effective immunotherapy. Curr. Opin. Immunol. 39, 44–51 (2016).
Kriegsman, B. A., Vangala, P., Chen, B. J., Meraner, P., Brass, A. L., Garber, M. et al. Frequent loss of IRF2 in cancers leads to immune evasion through decreased MHC class I antigen presentation and increased PD-L1 expression. J. Immunol. 203, 1999–2010 (2019).
Mariathasan, S., Turley, S. J., Nickles, D., Castiglioni, A., Yuen, K., Wang, Y. et al. TGFbeta attenuates tumour response to PD-L1 blockade by contributing to exclusion of T cells. Nature 554, 544–548 (2018).
Pardoll, D. M. The blockade of immune checkpoints in cancer immunotherapy. Nat. Rev. Cancer 12, 252–264 (2012).
Keenan, T. E., Burke, K. P. & Van Allen, E. M. Genomic correlates of response to immune checkpoint blockade. Nat. Med. 25, 389–402 (2019).
Nadal, E., Massuti, B., Domine, M., Garcia-Campelo, R., Cobo, M. & Felip, E. Immunotherapy with checkpoint inhibitors in non-small cell lung cancer: insights from long-term survivors. Cancer Immunol. Immunother. 68, 341–352 (2019).
Hermann-Kleiter, N., Klepsch, V., Wallner, S., Siegmund, K., Klepsch, S., Tuzlak, S. et al. The nuclear orphan receptor NR2F6 is a central checkpoint for cancer immune surveillance. Cell Rep. 12, 2072–2085 (2015).
Kvistborg, P., Philips, D., Kelderman, S., Hageman, L., Ottensmeier, C., Joseph-Pietras, D. et al. Anti-CTLA-4 therapy broadens the melanoma-reactive CD8+ T cell response. Sci. Transl. Med. 6, 254ra128 (2014).
Yost, K. E., Satpathy, A. T., Wells, D. K., Qi, Y., Wang, C., Kageyama, R. et al. Clonal replacement of tumor-specific T cells following PD-1 blockade. Nat. Med. 25, 1251–1259 (2019).
Hamid, O., Robert, C., Daud, A., Hodi, F. S., Hwu, W. J., Kefford, R. et al. Five-year survival outcomes for patients with advanced melanoma treated with pembrolizumab in KEYNOTE-001. Ann. Oncol. 30, 582–588 (2019).
Paz-Ares, L., Luft, A., Vicente, D., Tafreshi, A., Gumus, M., Mazieres, J. et al. Pembrolizumab plus chemotherapy for squamous non-small-cell lung cancer. N. Engl. J. Med. 379, 2040–2051 (2018).
Gandhi, L., Rodriguez-Abreu, D., Gadgeel, S., Esteban, E., Felip, E., De Angelis, F. et al. Pembrolizumab plus chemotherapy in metastatic non-small-cell lung cancer. N. Engl. J. Med. 378, 2078–2092 (2018).
de Miguel M. & Calvo, E. Clinical challenges of immune checkpoint inhibitors. Cancer Cell 38, 326–333 (2020).
Van Allen, E. M., Miao, D., Schilling, B., Shukla, S. A., Blank, C., Zimmer, L. et al. Genomic correlates of response to CTLA-4 blockade in metastatic melanoma. Science 350, 207–211 (2015).
Eroglu, Z., Zaretsky, J. M., Hu-Lieskovan, S., Kim, D. W., Algazi, A., Johnson, D. B. et al. High response rate to PD-1 blockade in desmoplastic melanomas. Nature 553, 347–350 (2018).
Hellmann, M. D., Ciuleanu, T. E., Pluzanski, A., Lee, J. S., Otterson, G. A., Audigier-Valette, C. et al. Nivolumab plus ipilimumab in lung cancer with a high tumor mutational burden. N. Engl. J. Med. 378, 2093–2104 (2018).
Havel, J. J., Chowell, D. & Chan, T. A. The evolving landscape of biomarkers for checkpoint inhibitor immunotherapy. Nat. Rev. Cancer 19, 133–150 (2019).
Samstein, R. M., Lee, C. H., Shoushtari, A. N., Hellmann, M. D., Shen, R., Janjigian, Y. Y. et al. Tumor mutational load predicts survival after immunotherapy across multiple cancer types. Nat. Genet. 51, 202–206 (2019).
Rizvi, N. A., Hellmann, M. D., Snyder, A., Kvistborg, P., Makarov, V., Havel, J. J. et al. Cancer immunology. Mutational landscape determines sensitivity to PD-1 blockade in non-small cell lung cancer. Science 348, 124–128 (2015).
Fancello, L., Gandini, S., Pelicci, P. G. & Mazzarella, L. Tumor mutational burden quantification from targeted gene panels: major advancements and challenges. J. Immunother. Cancer 7, 183 (2019).
Chan, T. A., Yarchoan, M., Jaffee, E., Swanton, C., Quezada, S. A., Stenzinger, A. et al. Development of tumor mutation burden as an immunotherapy biomarker: utility for the oncology clinic. Ann. Oncol. 30, 44–56 (2019).
Rooney, M. S., Shukla, S. A., Wu, C. J., Getz, G. & Hacohen, N. Molecular and genetic properties of tumors associated with local immune cytolytic activity. Cell 160, 48–61 (2015).
Yadav, M., Jhunjhunwala, S., Phung, Q. T., Lupardus, P., Tanguay, J., Bumbaca, S. et al. Predicting immunogenic tumour mutations by combining mass spectrometry and exome sequencing. Nature 515, 572–576 (2014).
Torti, D. & Trusolino, L. Oncogene addiction as a foundational rationale for targeted anti-cancer therapy: promises and perils. EMBO Mol. Med. 3, 623–636 (2011).
Weinstein, I. B. & Joe, A. Oncogene addiction. Cancer Res. 68, 3077–3080 (2008).
Sharma, P., Hu-Lieskovan, S., Wargo, J. A. & Ribas, A. Primary, adaptive, and acquired resistance to cancer immunotherapy. Cell 168, 707–723 (2017).
Zaretsky, J. M., Garcia-Diaz, A., Shin, D. S., Escuin-Ordinas, H., Hugo, W., Hu-Lieskovan, S. et al. Mutations associated with acquired resistance to PD-1 blockade in melanoma. N. Engl. J. Med. 375, 819–829 (2016).
Borghaei, H., Paz-Ares, L., Horn, L., Spigel, D. R., Steins, M., Ready, N. E. et al. Nivolumab versus docetaxel in advanced nonsquamous non-small-cell lung cancer. N. Engl. J. Med. 373, 1627–1639 (2015).
Garon, E. B., Rizvi, N. A., Hui, R., Leighl, N., Balmanoukian, A. S., Eder, J. P. et al. Pembrolizumab for the treatment of non-small-cell lung cancer. N. Engl. J. Med. 372, 2018–2028 (2015).
Reck, M., Rodriguez-Abreu, D., Robinson, A. G., Hui, R., Csoszi, T., Fulop, A. et al. Pembrolizumab versus chemotherapy for PD-L1-positive non-small-cell lung cancer. N. Engl. J. Med. 375, 1823–1833 (2016).
Martins, F., Sofiya, L., Sykiotis, G. P., Lamine, F., Maillard, M., Fraga, M. et al. Adverse effects of immune-checkpoint inhibitors: epidemiology, management and surveillance. Nat. Rev. Clin. Oncol. 16, 563–580 (2019).
Berner, F., Bomze, D., Diem, S., Ali, O. H., Fassler, M., Ring, S. et al. Association of checkpoint inhibitor-induced toxic effects with shared cancer and tissue antigens in non-small cell lung cancer. JAMA Oncol. 5, 1043–1047 (2019).
Andrews, A. Treating with checkpoint inhibitors-figure $1 million per patient. Am. Health Drug Benefits 8, 9 (2015).
Verma, V., Sprave, T., Haque, W., Simone, C. B. 2nd, Chang, J. Y., Welsh, J. W. et al. A systematic review of the cost and cost-effectiveness studies of immune checkpoint inhibitors. J. Immunother. Cancer 6, 128 (2018).
Tumeh, P. C., Harview, C. L., Yearley, J. H., Shintaku, I. P., Taylor, E. J., Robert, L. et al. PD-1 blockade induces responses by inhibiting adaptive immune resistance. Nature 515, 568–571 (2014).
Reck, M., Rodriguez-Abreu, D., Robinson, A. G., Hui, R., Csoszi, T., Fulop, A. et al. Updated analysis of KEYNOTE-024: pembrolizumab versus platinum-based chemotherapy for advanced non-small-cell lung cancer with PD-L1 tumor proportion score of 50% or greater. J. Clin. Oncol. 37, 537–546 (2019).
Gibney, G. T., Weiner, L. M. & Atkins, M. B. Predictive biomarkers for checkpoint inhibitor-based immunotherapy. Lancet Oncol. 17, e542–e551 (2016).
Meng, X., Huang, Z., Teng, F., Xing, L. & Yu, J. Predictive biomarkers in PD-1/PD-L1 checkpoint blockade immunotherapy. Cancer Treat. Rev. 41, 868–876 (2015).
Sivan, A., Corrales, L., Hubert, N., Williams, J. B., Aquino-Michaels, K., Earley, Z. M. et al. Commensal Bifidobacterium promotes antitumor immunity and facilitates anti-PD-L1 efficacy. Science 350, 1084–1089 (2015).
Vetizou, M., Pitt, J. M., Daillere, R., Lepage, P., Waldschmitt, N., Flament, C. et al. Anticancer immunotherapy by CTLA-4 blockade relies on the gut microbiota. Science 350, 1079–1084 (2015).
Derosa, L., Hellmann, M. D., Spaziano, M., Halpenny, D., Fidelle, M., Rizvi, H. et al. Negative association of antibiotics on clinical activity of immune checkpoint inhibitors in patients with advanced renal cell and non-small-cell lung cancer. Ann. Oncol. 29, 1437–1444 (2018).
Schett, A., Rothschild, S. I., Curioni-Fontecedro, A., Krahenbuhl, S., Fruh, M., Schmid, S. et al. Predictive impact of antibiotics in patients with advanced non small-cell lung cancer receiving immune checkpoint inhibitors: antibiotics immune checkpoint inhibitors in advanced NSCLC. Cancer Chemother. Pharmacol. 85, 121–131 (2020).
Pinato, D. J., Gramenitskaya, D., Altmann, D. M., Boyton, R. J., Mullish, B. H., Marchesi, J. R. et al. Antibiotic therapy and outcome from immune-checkpoint inhibitors. J. Immunother. Cancer 7, 287 (2019).
Chalabi, M., Cardona, A., Nagarkar, D. R., Dhawahir Scala, A., Gandara, D. R., Rittmeyer, A. et al. Efficacy of chemotherapy and atezolizumab in patients with non-small-cell lung cancer receiving antibiotics and proton pump inhibitors: pooled post hoc analyses of the OAK and POPLAR trials. Ann. Oncol. 31, 525–531 (2020).
Pinato, D. J., Howlett, S., Ottaviani, D., Urus, H., Patel, A., Mineo, T. et al. Association of prior antibiotic treatment with survival and response to immune checkpoint inhibitor therapy in patients with cancer. JAMA Oncol. 5, 1774–1778 (2019).
Routy, B., Le Chatelier, E., Derosa, L., Duong, C. P. M., Alou, M. T., Daillere, R. et al. Gut microbiome influences efficacy of PD-1-based immunotherapy against epithelial tumors. Science 359, 91–97 (2018).
Fessler, J., Matson, V. & Gajewski, T. F. Exploring the emerging role of the microbiome in cancer immunotherapy. J. Immunother. Cancer 7, 108 (2019).
Balachandran, V. P., Luksza, M., Zhao, J. N., Makarov, V., Moral, J. A., Remark, R. et al. Identification of unique neoantigen qualities in long-term survivors of pancreatic cancer. Nature 551, 512–516 (2017).
Sioud, M. T-cell cross-reactivity may explain the large variation in how cancer patients respond to checkpoint inhibitors. Scand. J. Immunol. https://doi.org/10.1111/sji.12643 (2018).
Sahin, U. & Tureci, O. Personalized vaccines for cancer immunotherapy. Science 359, 1355–1360 (2018).
Leng, Q., Tarbe, M., Long, Q. & Wang, F. Pre-existing heterologous T-cell immunity and neoantigen immunogenicity. Clin. Transl. Immunol. 9, e01111 (2020).
Finlay, B. B., Goldszmid, R., Honda, K., Trinchieri, G., Wargo, J. & Zitvogel, L. Can we harness the microbiota to enhance the efficacy of cancer immunotherapy? Nat. Rev. Immunol. 20, 522–528 (2020).
Shi, Y., Zheng, W., Yang, K., Harris, K. G., Ni, K., Xue, L. et al. Intratumoral accumulation of gut microbiota facilitates CD47-based immunotherapy via STING signaling. J. Exp. Med. 217, e20192282 (2020).
Nejman, D., Livyatan, I., Fuks, G., Gavert, N., Zwang, Y., Geller, L. T. et al. The human tumor microbiome is composed of tumor type-specific intracellular bacteria. Science 368, 973–980 (2020).
Man, W. H., de Steenhuijsen Piters, W. A. & Bogaert, D. The microbiota of the respiratory tract: gatekeeper to respiratory health. Nat. Rev. Microbiol. 15, 259–270 (2017).
Wypych, T. P., Wickramasinghe, L. C. & Marsland, B. J. The influence of the microbiome on respiratory health. Nat. Immunol. 20, 1279–1290 (2019).
Ravel, J., Gajer, P., Abdo, Z., Schneider, G. M., Koenig, S. S., McCulle, S. L. et al. Vaginal microbiome of reproductive-age women. Proc. Natl Acad. Sci. USA 108, 4680–4687 (2011).
Rawls, M. & Ellis, A. K. The microbiome of the nose. Ann. Allergy Asthma Immunol. 122, 17–24 (2019).
Kolenbrander, P. E. Oral microbial communities: biofilms, interactions, and genetic systems. Annu. Rev. Microbiol. 54, 413–437 (2000).
Wieczorek, M., Abualrous, E. T., Sticht, J., Alvaro-Benito, M., Stolzenberg, S., Noe, F. et al. Major histocompatibility complex (MHC) class I and MHC class II proteins: conformational plasticity in antigen presentation. Front. Immunol. 8, 292 (2017).
Stryhn, A., Pedersen, L. O., Holm, A. & Buus, S. Longer peptide can be accommodated in the MHC class I binding site by a protrusion mechanism. Eur. J. Immunol. 30, 3089–3099 (2000).
Amorim Franco, T. M. & Blanchard, J. S. Bacterial branched-chain amino acid biosynthesis: structures, mechanisms, and drugability. Biochemistry 56, 5849–5865 (2017).
Tack, D. S., Cole, A. C., Shroff, R., Morrow, B. R. & Ellington, A. D. Evolving bacterial fitness with an expanded genetic code. Sci. Rep. 8, 3288 (2018).
Thursby, E. & Juge, N. Introduction to the human gut microbiota. Biochem. J. 474, 1823–1836 (2017).
Steers, N. J., Currier, J. R., Jobe, O., Tovanabutra, S., Ratto-Kim, S., Marovich, M. A. et al. Designing the epitope flanking regions for optimal generation of CTL epitopes. Vaccine 32, 3509–3516 (2014).
Textoris-Taube, K., Henklein, P., Pollmann, S., Bergann, T., Weisshoff, H., Seifert, U. et al. The N-terminal flanking region of the TRP2360-368 melanoma antigen determines proteasome activator PA28 requirement for epitope liberation. J. Biol. Chem. 282, 12749–12754 (2007).
Spencer, J. & Sollid, L. M. The human intestinal B-cell response. Mucosal Immunol. 9, 1113–1124 (2016).
Okai, S., Usui, F., Yokota, S., Hori, I. Y., Hasegawa, M., Nakamura, T. et al. High-affinity monoclonal IgA regulates gut microbiota and prevents colitis in mice. Nat. Microbiol. 1, 16103 (2016).
Chang, S. Y., Song, J. H., Guleng, B., Cotoner, C. A., Arihiro, S., Zhao, Y. et al. Circulatory antigen processing by mucosal dendritic cells controls CD8(+) T cell activation. Immunity 38, 153–165 (2013).
Persson, E. K., Scott, C. L., Mowat, A. M. & Agace, W. W. Dendritic cell subsets in the intestinal lamina propria: ontogeny and function. Eur. J. Immunol. 43, 3098–3107 (2013).
Feng, T. & Elson, C. O. Adaptive immunity in the host-microbiota dialog. Mucosal Immunol. 4, 15–21 (2011).
Russler-Germain, E. V., Rengarajan, S. & Hsieh, C. S. Antigen-specific regulatory T-cell responses to intestinal microbiota. Mucosal Immunol. 10, 1375–1386 (2017).
Chinthrajah, R. S., Hernandez, J. D., Boyd, S. D., Galli, S. J. & Nadeau, K. C. Molecular and cellular mechanisms of food allergy and food tolerance. J. Allergy Clin. Immunol. 137, 984–997 (2016).
Tordesillas, L. & Berin, M. C. Mechanisms of oral tolerance. Clin. Rev. Allergy Immunol. 55, 107–117 (2018).
Nomura, M., Nagatomo, R., Doi, K., Shimizu, J., Baba, K., Saito, T. et al. Association of short-chain fatty acids in the gut microbiome with clinical response to treatment with nivolumab or pembrolizumab in patients with solid cancer tumors. JAMA Netw. Open 3, e202895 (2020).
Malczewski, A. B., Navarro, S., Coward, J. I. & Ketheesan, N. Microbiome-derived metabolome as a potential predictor of response to cancer immunotherapy. J. Immunother. Cancer 8, e001383 (2020).
Mager, L. F., Burkhard, R., Pett, N., Cooke, N. C. A., Brown, K., Ramay, H. et al. Microbiome-derived inosine modulates response to checkpoint inhibitor immunotherapy. Science 369, 1481–1489 (2020).
Staley, C., Kaiser, T., Beura, L. K., Hamilton, M. J., Weingarden, A. R., Bobr, A. et al. Stable engraftment of human microbiota into mice with a single oral gavage following antibiotic conditioning. Microbiome 5, 87 (2017).
Buque, A. & Galluzzi, L. Modeling tumor immunology and immunotherapy in mice. Trends Cancer. 4, 599–601 (2018).
Gargiulo, G. Next-generation in vivo modeling of human cancers. Front. Oncol. 8, 429 (2018).
Hoffman, R. M. Patient-derived orthotopic xenografts: better mimic of metastasis than subcutaneous xenografts. Nat. Rev. Cancer 15, 451–452 (2015).
Rongvaux, A., Willinger, T., Martinek, J., Strowig, T., Gearty, S. V., Teichmann, L. L. et al. Development and function of human innate immune cells in a humanized mouse model. Nat. Biotechnol. 32, 364–372 (2014).
Theocharides, A. P., Rongvaux, A., Fritsch, K., Flavell, R. A. & Manz, M. G. Humanized hemato-lymphoid system mice. Haematologica 101, 5–19 (2016).
Hermans, I. F., Daish, A., Yang, J., Ritchie, D. S. & Ronchese, F. Antigen expressed on tumor cells fails to elicit an immune response, even in the presence of increased numbers of tumor-specific cytotoxic T lymphocyte precursors. Cancer Res. 58, 3909–3917 (1998).
Prevost-Blondel, A., Zimmermann, C., Stemmer, C., Kulmburg, P., Rosenthal, F. M. & Pircher, H. Tumor-infiltrating lymphocytes exhibiting high ex vivo cytolytic activity fail to prevent murine melanoma tumor growth in vivo. J. Immunol. 161, 2187–2194 (1998).
Cheng, H. W., Onder, L., Cupovic, J., Boesch, M., Novkovic, M., Pikor, N. et al. CCL19-producing fibroblastic stromal cells restrain lung carcinoma growth by promoting local antitumor T-cell responses. J. Allergy Clin. Immunol. 142, 1257–1271 e4 (2018).
Moore, M. W., Carbone, F. R. & Bevan, M. J. Introduction of soluble protein into the class I pathway of antigen processing and presentation. Cell 54, 777–785 (1988).
Bellone, M., Cantarella, D., Castiglioni, P., Crosti, M. C., Ronchetti, A., Moro, M. et al. Relevance of the tumor antigen in the validation of three vaccination strategies for melanoma. J. Immunol. 165, 2651–2656 (2000).
Weigelin, B., Bolanos, E., Teijeira, A., Martinez-Forero, I., Labiano, S., Azpilikueta, A. et al. Focusing and sustaining the antitumor CTL effector killer response by agonist anti-CD137 mAb. Proc. Natl Acad. Sci. USA 112, 7551–7556 (2015).
Kurtulus, S. & Hildeman, D. Assessment of CD4(+) and CD8 (+) T cell responses using MHC class I and II tetramers. Methods Mol. Biol. 979, 71–79 (2013).
Estcourt, M. J., Ramsay, A. J., Brooks, A., Thomson, S. A., Medveckzy, C. J. & Ramshaw, I. A. Prime-boost immunization generates a high frequency, high-avidity CD8(+) cytotoxic T lymphocyte population. Int. Immunol. 14, 31–37 (2002).
Janelle, V., Langlois, M. P., Lapierre, P., Charpentier, T., Poliquin, L. & Lamarre, A. The strength of the T cell response against a surrogate tumor antigen induced by oncolytic VSV therapy does not correlate with tumor control. Mol. Ther. 22, 1198–1210 (2014).
van Vloten, J. P., Santry, L. A., McAusland, T. M., Karimi, K., McFadden, G., Petrik, J. J. et al. Quantifying antigen-specific T cell responses when using antigen-agnostic immunotherapies. Mol. Ther. Methods Clin. Dev. 13, 154–166 (2019).
Kemp, C. J. Animal models of chemical carcinogenesis: driving breakthroughs in cancer research for 100 years. Cold Spring Harb. Protoc. 2015, 865–874 (2015).
Jackson, E. L., Willis, N., Mercer, K., Bronson, R. T., Crowley, D., Montoya, R. et al. Analysis of lung tumor initiation and progression using conditional expression of oncogenic K-ras. Genes Dev. 15, 3243–3248 (2001).
Singh, M., Lima, A., Molina, R., Hamilton, P., Clermont, A. C., Devasthali, V. et al. Assessing therapeutic responses in Kras mutant cancers using genetically engineered mouse models. Nat. Biotechnol. 28, 585–593 (2010).
Guy, C. T., Cardiff, R. D. & Muller, W. J. Induction of mammary tumors by expression of polyomavirus middle T oncogene: a transgenic mouse model for metastatic disease. Mol. Cell. Biol. 12, 954–961 (1992).
Qiu, T. H., Chandramouli, G. V., Hunter, K. W., Alkharouf, N. W., Green, J. E. & Liu, E. T. Global expression profiling identifies signatures of tumor virulence in MMTV-PyMT-transgenic mice: correlation to human disease. Cancer Res. 64, 5973–5981 (2004).
Guy, C. T., Webster, M. A., Schaller, M., Parsons, T. J., Cardiff, R. D. & Muller, W. J. Expression of the neu protooncogene in the mammary epithelium of transgenic mice induces metastatic disease. Proc. Natl Acad. Sci. USA 89, 10578–10582 (1992).
Aguirre, A. J., Bardeesy, N., Sinha, M., Lopez, L., Tuveson, D. A., Horner, J. et al. Activated Kras and Ink4a/Arf deficiency cooperate to produce metastatic pancreatic ductal adenocarcinoma. Genes Dev. 17, 3112–3126 (2003).
Dunn, G. P., Old, L. J. & Schreiber, R. D. The three Es of cancer immunoediting. Annu. Rev. Immunol. 22, 329–360 (2004).
Zitvogel, L., Tesniere, A. & Kroemer, G. Cancer despite immunosurveillance: immunoselection and immunosubversion. Nat. Rev. Immunol. 6, 715–727 (2006).
Chung, W. J., Daemen, A., Cheng, J. H., Long, J. E., Cooper, J. E., Wang, B. E. et al. Kras mutant genetically engineered mouse models of human cancers are genomically heterogeneous. Proc. Natl Acad. Sci. USA 114, E10947–E10955 (2017).
Reynisson, B., Alvarez, B., Paul, S., Peters, B. & Nielsen, M. NetMHCpan-4.1 and NetMHCIIpan-4.0: improved predictions of MHC antigen presentation by concurrent motif deconvolution and integration of MS MHC eluted ligand data. Nucleic Acids Res. 48, W449–W454 (2020).
Boesch, M., Cosma, A. & Sopper, S. Flow cytometry: to dump or not to dump. J. Immunol. 201, 1813–1815 (2018).
Boesch, M., Onder, L., Cheng, H. W., Novkovic, M., Morbe, U., Sopper, S. et al. Interleukin 7-expressing fibroblasts promote breast cancer growth through sustenance of tumor cell stemness. Oncoimmunology 7, e1414129 (2018).
Davis, M. M., Altman, J. D. & Newell, E. W. Interrogating the repertoire: broadening the scope of peptide-MHC multimer analysis. Nat. Rev. Immunol. 11, 551–558 (2011).
Ott, P. A., Hu, Z., Keskin, D. B., Shukla, S. A., Sun, J., Bozym, D. J. et al. An immunogenic personal neoantigen vaccine for patients with melanoma. Nature 547, 217–221 (2017).
Keskin, D. B., Anandappa, A. J., Sun, J., Tirosh, I., Mathewson, N. D., Li, S. et al. Neoantigen vaccine generates intratumoral T cell responses in phase Ib glioblastoma trial. Nature 565, 234–239 (2019).
Linnemann, C., Mezzadra, R. & Schumacher, T. N. TCR repertoires of intratumoral T-cell subsets. Immunol. Rev. 257, 72–82 (2014).
Boesch, M., Zeimet, A. G., Reimer, D., Schmidt, S., Gastl, G., Parson, W. et al. The side population of ovarian cancer cells defines a heterogeneous compartment exhibiting stem cell characteristics. Oncotarget 5, 7027–7039 (2014).
Matson, V., Fessler, J., Bao, R., Chongsuwat, T., Zha, Y., Alegre, M. L. et al. The commensal microbiome is associated with anti-PD-1 efficacy in metastatic melanoma patients. Science 359, 104–108 (2018).
Korem, T., Zeevi, D., Suez, J., Weinberger, A., Avnit-Sagi, T., Pompan-Lotan, M. et al. Growth dynamics of gut microbiota in health and disease inferred from single metagenomic samples. Science 349, 1101–1106 (2015).
Sekirov, I., Russell, S. L., Antunes, L. C. & Finlay, B. B. Gut microbiota in health and disease. Physiol. Rev. 90, 859–904 (2010).
Botticelli, A., Vernocchi, P., Marini, F., Quagliariello, A., Cerbelli, B., Reddel, S. et al. Gut metabolomics profiling of non-small cell lung cancer (NSCLC) patients under immunotherapy treatment. J. Transl. Med. 18, 49 (2020).
Song, P., Yang, D., Wang, H., Cui, X., Si, X., Zhang, X. et al. Relationship between intestinal flora structure and metabolite analysis and immunotherapy efficacy in Chinese NSCLC patients. Thorac. Cancer 11, 1621–1632 (2020).
Mohiuddin, J. J., Chu, B., Facciabene, A., Poirier, K., Wang, X., Doucette, A. et al. Association of antibiotic exposure with survival and toxicity in patients with melanoma receiving immunotherapy. J. Natl Cancer Inst. 113, 162–170 (2020).
Riquelme, E., Zhang, Y., Zhang, L., Montiel, M., Zoltan, M., Dong, W. et al. Tumor microbiome diversity and composition influence pancreatic cancer outcomes. Cell 178, 795.e12–806.e12 (2019).
Jin, Y., Dong, H., Xia, L., Yang, Y., Zhu, Y., Shen, Y. et al. The diversity of gut microbiome is associated with favorable responses to anti-programmed death 1 immunotherapy in Chinese patients with NSCLC. J. Thorac. Oncol. 14, 1378–1389 (2019).
Khanna, S., Vazquez-Baeza, Y., Gonzalez, A., Weiss, S., Schmidt, B., Muniz-Pedrogo, D. A. et al. Changes in microbial ecology after fecal microbiota transplantation for recurrent C. difficile infection affected by underlying inflammatory bowel disease. Microbiome 5, 55 (2017).
Vaughn, B. P., Vatanen, T., Allegretti, J. R., Bai, A., Xavier, R. J., Korzenik, J. et al. Increased intestinal microbial diversity following fecal microbiota transplant for active Crohn’s disease. Inflamm. Bowel Dis. 22, 2182–2190 (2016).
Mittal, D., Gubin, M. M., Schreiber, R. D. & Smyth, M. J. New insights into cancer immunoediting and its three component phases–elimination, equilibrium and escape. Curr. Opin. Immunol. 27, 16–25 (2014).
So, T., Takenoyama, M., Mizukami, M., Ichiki, Y., Sugaya, M., Hanagiri, T. et al. Haplotype loss of HLA class I antigen as an escape mechanism from immune attack in lung cancer. Cancer Res. 65, 5945–5952 (2005).
Leone, P., Shin, E. C., Perosa, F., Vacca, A., Dammacco, F. & Racanelli, V. MHC class I antigen processing and presenting machinery: organization, function, and defects in tumor cells. J. Natl Cancer Inst. 105, 1172–1187 (2013).
Montesion, M., Murugesan, K., Jin, D. X., Sharaf, R., Sanchez, N., Guria, A. et al. Somatic HLA class I loss is a widespread mechanism of immune evasion which refines the use of tumor mutational burden as a biomarker of checkpoint inhibitor response. Cancer Discov. 11, 282–292 (2020).
Author information
Authors and Affiliations
Contributions
Wrote the first draft of the paper: M.B., F.B., M.H.B. Generated the figures: M.B. Wrote the final version of the paper: M.B., F.B., S.I.R., M.T., M.J., M.F., M.H.B. Approved the paper for publication: M.B., F.B., S.I.R., M.T., M.J., M.F., M.H.B.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Data availability
Not applicable.
Competing interests
M.B. serves as an advisor for Pantec Biosolutions AG. The other authors have no potential conflicts of interest to declare. No medical writer or other non-author was involved in the preparation of the manuscript.
Funding information
This work was supported by the Lungenliga St. Gallen-Appenzell (grant to M.B., no grant number available).
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Boesch, M., Baty, F., Rothschild, S.I. et al. Tumour neoantigen mimicry by microbial species in cancer immunotherapy. Br J Cancer 125, 313–323 (2021). https://doi.org/10.1038/s41416-021-01365-2
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41416-021-01365-2
This article is cited by
-
Leveraging circulating microbiome signatures to predict tumor immune microenvironment and prognosis of patients with non-small cell lung cancer
Journal of Translational Medicine (2023)
-
Microbiota as the unifying factor behind the hallmarks of cancer
Journal of Cancer Research and Clinical Oncology (2023)
