
Abstract
Group 2 innate lymphoid cells (ILC2s) regulate inflammation and immunity in mammalian tissues1,2. Although ILC2s are found in cancers of these tissues3, their roles in cancer immunity and immunotherapy are unclear. Here we show that ILC2s infiltrate pancreatic ductal adenocarcinomas (PDACs) to activate tissue-specific tumour immunity. Interleukin-33 (IL33) activates tumour ILC2s (TILC2s) and CD8+ T cells in orthotopic pancreatic tumours but not heterotopic skin tumours in mice to restrict pancreas-specific tumour growth. Resting and activated TILC2s express the inhibitory checkpoint receptor PD-1. Antibody-mediated PD-1 blockade relieves ILC2 cell-intrinsic PD-1 inhibition to expand TILC2s, augment anti-tumour immunity, and enhance tumour control, identifying activated TILC2s as targets of anti-PD-1 immunotherapy. Finally, both PD-1+ TILC2s and PD-1+ T cells are present in most human PDACs. Our results identify ILC2s as anti-cancer immune cells for PDAC immunotherapy. More broadly, ILC2s emerge as tissue-specific enhancers of cancer immunity that amplify the efficacy of anti-PD-1 immunotherapy. As ILC2s and T cells co-exist in human cancers and share stimulatory and inhibitory pathways, immunotherapeutic strategies to collectively target anti-cancer ILC2s and T cells may be broadly applicable.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 51 print issues and online access
$199.00 per year
only $3.90 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout




Similar content being viewed by others

Data availability
Source code for immune quantification is available in Supplementary Data 1. Bulk RNA-seq data are available under Gene Expression Omnibus (GEO) accession number GSE129388. scRNA-seq data are available under GEO accession number GSE136720. Source data are provided for all experiments. All other data are available from the corresponding author upon reasonable request.
References
Vivier, E., van de Pavert, S. A., Cooper, M. D. & Belz, G. T. The evolution of innate lymphoid cells. Nat. Immunol. 21, 790–794 (2016).
Vivier, E. et al. Innate lymphoid cells: 10 years on. Cell 174, 1054–1066 (2018).
Salimi, M. et al. Activated innate lymphoid cell populations accumulate in human tumour tissues. BMC Cancer 18, 341 (2018).
Balachandran, V. P. et al. Identification of unique neoantigen qualities in long-term survivors of pancreatic cancer. Nature 551, 512–516 (2017).
Hingorani, S. R. et al. Trp53R172H and KrasG12D cooperate to promote chromosomal instability and widely metastatic pancreatic ductal adenocarcinoma in mice. Cancer Cell 7, 469–483 (2005).
Pylayeva-Gupta, Y., Lee, K. E., Hajdu, C. H., Miller, G. & Bar-Sagi, D. Oncogenic Kras-induced GM-CSF production promotes the development of pancreatic neoplasia. Cancer Cell 21, 836–847 (2012).
Li, J. et al. Tumor cell-intrinsic factors underlie heterogeneity of immune cell infiltration and response to immunotherapy. Immunity 49, 178–193.e7 (2018).
Brestoff, J. R. et al. Group 2 innate lymphoid cells promote beiging of white adipose tissue and limit obesity. Nature 519, 242–246 (2015).
Gasteiger, G., Fan, X., Dikiy, S., Lee, S. Y. & Rudensky, A. Y. Tissue residency of innate lymphoid cells in lymphoid and nonlymphoid organs. Science 350, 981–985 (2015).
Kirchberger, S. et al. Innate lymphoid cells sustain colon cancer through production of interleukin-22 in a mouse model. J. Exp. Med. 210, 917–931 (2013).
Monticelli, L. A. et al. Arginase 1 is an innate lymphoid-cell-intrinsic metabolic checkpoint controlling type 2 inflammation. Nat. Immunol. 17, 656–665 (2016).
Neesse, A. et al. CTGF antagonism with mAb FG-3019 enhances chemotherapy response without increasing drug delivery in murine ductal pancreas cancer. Proc. Natl Acad. Sci. USA 110, 12325–12330 (2013).
Hardman, C. S., Panova, V. & McKenzie, A. N. J. IL-33 citrine reporter mice reveal the temporal and spatial expression of IL-33 during allergic lung inflammation. Eur. J. Immunol. 43, 488–498 (2013).
Talabot-Ayer, D. et al. The mouse interleukin (Il)33 gene is expressed in a cell type- and stimulus-dependent manner from two alternative promoters. J. Leukoc. Biol. 91, 119–125 (2012).
Ricardo-Gonzalez, R. R. et al. Tissue signals imprint ILC2 identity with anticipatory function. Nat. Immunol. 19, 1093–1099 (2018).
Dalmas, E. et al. Interleukin-33-activated islet-resident innate lymphoid cells promote insulin secretion through myeloid cell retinoic acid production. Immunity 47, 928–942.e7 (2017).
Oliphant, C. J. et al. MHCII-mediated dialog between group 2 innate lymphoid cells and CD4+ T cells potentiates type 2 immunity and promotes parasitic helminth expulsion. Immunity 41, 283–295 (2014).
Böttcher, J. P. et al. NK cells stimulate recruitment of cDC1 into the tumor microenvironment promoting cancer immune control. Cell 172, 1022–1037.e14 (2018).
Yu, Y. et al. Single-cell RNA-seq identifies a PD-1hi ILC progenitor and defines its development pathway. Nature 539, 102–106 (2016).
Taylor, S. et al. PD-1 regulates KLRG1+ group 2 innate lymphoid cells. J. Exp. Med. 214, 1663–1678 (2017).
Kim, J. et al. Intratumorally establishing type 2 innate lymphoid cells blocks tumor growth. J. Immunol. 196, 2410–2423 (2016).
Diana, A. et al. Prognostic value, localization and correlation of PD-1/PD-L1, CD8 and FOXP3 with the desmoplastic stroma in pancreatic ductal adenocarcinoma. Oncotarget 7, 40992–41004 (2016).
Donovan, C. et al. Roles for T/B lymphocytes and ILC2s in experimental chronic obstructive pulmonary disease. J. Leukoc. Biol. 105, 143–150 (2019).
Evans, R. A. et al. Lack of immunoediting in murine pancreatic cancer reversed with neoantigen. JCI Insight 1, 88328 (2016).
Sastra, S. A. & Olive, K. P. Quantification of murine pancreatic tumors by high-resolution ultrasound. Methods Mol. Biol. 980, 249–266 (2013).
Monticelli, L. A. et al. Innate lymphoid cells promote lung-tissue homeostasis after infection with influenza virus. Nat. Immunol. 12, 1045–1054 (2011).
Ma, Z. et al. Augmentation of immune checkpoint cancer immunotherapy with IL18. Clin. Cancer Res. 22, 2969–2980 (2016).
Rooney, M. S., Shukla, S. A., Wu, C. J., Getz, G. & Hacohen, N. Molecular and genetic properties of tumors associated with local immune cytolytic activity. Cell 160, 48–61 (2015).
Yarilin, D. et al. Machine-based method for multiplex in situ molecular characterization of tissues by immunofluorescence detection. Sci. Rep. 5, 9534 (2015).
Butler, A., Hoffman, P., Smibert, P., Papalexi, E. & Satija, R. Integrating single-cell transcriptomic data across different conditions, technologies, and species. Nat. Biotechnol. 36, 411–420 (2018).
Acknowledgements
We thank J. Novak, J. Moore and E. Patterson for editorial assistance; B. Medina, G. Vitiello, J. Zhang, S. Zeng, F. Rossi, J. Loo, N. Param, J. Maltbaek, O. Grbovic-Huezo, Y. Senbabaoglu, M. Gigoux, R. Giese and S. Budhu for helpful discussions and technical assistance; and the Epigenomics Core of Weill Cornell Medical College for technical assistance with scRNA-seq. This work was supported by the V Foundation Convergence Scholar Grant (J.A.M., J.D.W., V.P.B.), the Stand Up to Cancer Convergence Award (J.D.W., V.P.B.), the National Cancer Institute K12CA184746-01A1 (V.P.B.), the Damon Runyon Clinical Investigator Award (V.P.B.), the Ben and Rose Cole Pria Foundation Scholar Award (V.P.B.), the Sarah Min and Matthew Pincus Pancreatic Cancer Immunotherapy Award (V.P.B.), an administrative supplement to NIH P30-CA008748 (S.D.L., V.P.B.), NIH R01 CA204228, NIH P30CA023108 (S.D.L.), Swim Across America and the Ludwig Institute for Cancer Research (J.D.W., T.M.), and the Parker Institute for Cancer Immunotherapy (J.D.W., T.M.). Services by the MSKCC Small-Animal Core Facility and Integrated Genomics Core were funded by the National Cancer Institute Cancer Center Support Grant (P30 CA008748-48), Cycle for Survival, and the Marie-Josée and Henry R. Kravis Center for Molecular Oncology.
Author information
Authors and Affiliations
Contributions
V.P.B. conceived the study. J.A.M, J.L., L.A.R., S.D.L., J.D.W., R.P.D., T.M. and V.P.B. designed all the experiments. J.A.M, J.L., L.A.R., J.R., J.Z., A.R. and V.P.B. performed all the experiments. B.G. assisted with generation of bone marrow chimaeras. J.L., Z.S. and D.R. analysed the scRNA-seq results. G.A. and U.B. performed the pathologic analyses. E.E., Y.P. and D.A.T. generated and assisted in experiments on autochthonous KPC mice. M. Gururajan provided technical assistance with the PD-1 blocking antibody. M. Gönen provided statistical oversight. J.A.M., J.L., L.A.R., J.R., S.D.L., R.P.D., T.M. and V.P.B. analysed all the data. All authors interpreted the data. J.A.M. and V.P.B. wrote the manuscript with input from all authors.
Corresponding authors
Ethics declarations
Competing interests
V.P.B. is a recipient of an immuno-oncology translational research grant from Bristol-Myers Squibb and is an inventor on a patent application related to work on neoantigen modelling. S.D.L. is a member of the scientific advisory board of Nybo Pharmaceuticals, and co-founder of Episteme Prognostics. J.D.W. is a consultant for Adaptive Biotech, Advaxis, Amgen, Apricity, Array BioPharma, Ascentage Pharma, Astellas, Bayer, Beigene, Bristol-Myers Squibb, Celgene, Chugai, Elucida, Eli Lilly, F Star, Genentech, Imvaq, Janssen, Kleo Pharma, Linneaus, MedImmune, Merck, Neon Therapuetics, Ono, Polaris Pharma, Polynoma, Psioxus, Puretech, Recepta, Trieza, Sellas Life Sciences, Serametrix, Surface Oncology and Syndax; is a recipient of research support from Bristol-Myers Squibb, Medimmune, Merck Pharmaceuticals and Genentech; has equity in Potenza Therapeutics, Tizona Pharmaceuticals, Adaptive Biotechnologies, Elucida, Imvaq, Beigene, Trieza and Linneaus; and has received an honorarium from Esanex. T.M. is a consultant for Immunos Therapeutics and Pfizer; is a co-founder with equity in Imvaq Therapeutics; receives research funding from Bristol-Myers Squibb, Surface Oncology, Kyn Therapeutics, Infinity Pharmaceuticals Inc., Peregrine Pharmaceuticals Inc., Adaptive Biotechnologies, Leap Therapeutics Inc. and Aprea; and is an inventor on patent applications related to work on Oncolytic Viral Therapy, Alpha Virus Based Vaccine, Neo Antigen Modeling, CD40, GITR, OX40, PD-1 and CTLA-4. M. Gururajan is an employee of Bristol-Myers Squibb and has financial interest in the company.
Additional information
Peer review information Nature thanks Richard Locksley and the other, anonymous, reviewer(s) for their contribution to the peer review of this work.
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data figures and tables
Extended Data Fig. 1 Identification of IL33-dependent ILCs in pancreatic cancer.
a, Gating strategy to identify human ILCs. The first plot was pre-gated on live (DRAQ7−) cells and singlets. Lineage 1 cocktail: CD5, CD11b, CD11c, CD16, FcεR1. Lineage 2 cocktail: CD3, CD19, TCRα/β. ILCs were identified as lineage− CD56− CD25+ CD127+ cells. FMO, fluorescence minus one. b, Representative image of immunofluorescence of ILC2s in tumour tissue microarrays from short- and long-term PDAC survivors (n = 96). Arrows, putative ILC2s. c, Top, overall survival of patients with more (high) or less (low) than the median intratumoral mRNA level of ILC-stimulating cytokines. Bottom, correlation between expression of ILC-activating cytokines and immune cytolytic index (CYT) in short- and long-term survivors of human PDAC. Curves were fit by linear regression. n = 25. d, Gating strategy to identify mouse ILCs. The first plot was pre-gated on live (DRAQ7−) cells and singlets. Lineage 1 cocktail: CD5, CD11b, CD11c, FcεR1. Lineage 2 cocktail: CD3, CD19. ILCs were identified as lineage− NK1.1− CD25+ CD127+, and ILC2s were identified as lineage− NK1.1− CD25+ ST2+ cells. Gating on orthotopic PDAC mice shown. e, Intratumoral ILC frequency in orthotopic PDAC mice established with KPC cell lines 8-1, 18-3, and in autochthonous KPC mice with spontaneous PDAC (KPCSpont). Composite ILC frequencies from Fig. 1d and others are included for comparison (KPC 4662). f, Phenotype of ILCs in PDAC mice. Grey curves, isotype controls; numbers, mean fluorescence intensity. g, Expansion kinetics of ILCs in tissues from PDAC mice. h, Changes in non-ILC cell frequency in Rag2−/− PDAC mice treated with anti-CD90.2 or isotype antibodies. Data were collected 14 days (d–f), 10 days (h), or at the indicated time points after tumour implantation. n indicates individual mice analysed separately in at least two independent experiments with n ≥ 2 per group. Median ± s.e.m; horizontal bars show median. P values determined by two-sided log-rank test (c, top), linear regression (c, bottom), or two-tailed Mann–Whitney test (g). P values in g indicate tumour comparisons to all other tissues.
Extended Data Fig. 2 Host-derived IL33 activates pancreatic ILC2s.
a, mRNA expression of ILC1- (IL12, IL15, IL18), ILC2- (IL25, IL33, TSLP), and ILC3-inducer cytokines (IL23) and the IL33 receptor (ST2) in orthotopic PDAC tumours (left) and autochthonous PDAC tumours in KPC mice from a previously published mRNA microarray (right)12. b, Representative IL33 immunohistochemistry (IHC) of IL33Low and IL33High human (tissue microarray, n = 96) and mouse PDAC (n = 3 per group). c, Frequency of patients with PDAC demonstrating IL33 positivity by IHC in a human PDAC tumour microarray. d, Multiplexed immunofluorescence for IL33, ductal marker CK19, and myeloid markers CD11b and Iba in mouse PDAC (top). Arrows, IL33-expressing cells. IL33 mean fluorescence intensity (MFI) in non-immune (CD45−), immune (CD45+), macrophage (TAM), and monocytic and granulocytic myeloid-derived suppressor cell (M-MDSC and G-MDSC) populations in tumours from IL33Cit reporter PDAC mice (bottom). e, Representative IL33 protein expression shown by IHC in orthotopic PDAC tumours in Il33+/+ (WT) mice, and non-tumour-bearing pancreata in Il33–/– mice (n = 3 per group). f, ILC frequency (top) and cell number (bottom) in organs and DLNs of Il33+/+ and Il33−/− orthotopic PDAC mice. g, Gating and frequency of IL4 and IL5 expression in intratumoral ILCs in Il33+/+ and Il33−/− orthotopic PDAC mice. h, i, ILC2 (h) and immune cell frequencies (i) in orthotopic Rag2–/– and Rag2–/–γc–/– PDAC mice with or without treatment with rIL33. j, Frequency of ST2+ tumour ILCs in mice with subcutaneous (SQ) and orthotopic PDACs. k, Tumours in orthotopic and subcutaneous PDAC mice. l, Tumour weight in Il33+/+ and Il33−/− littermate PDAC mice. m, Experimental schema of bone-marrow chimaeras to evaluate contribution of haematopoietic cell-derived IL33 to tumour control. n, o, Haematopoietic cell reconstitution (n) and tumour weight (o) in irradiated CD45.1 congenic mice reconstituted with either CD45.2 Il33+/+ or CD45.2 Il33−/− bone marrow. Data were collected 14 (a, b, d-g, j, k (orthotopic), l, o), 28 (k (subcutaneous)), or 10 (h, i) days after tumour implantation. Horizontal bars show median. n indicates individual mice analysed separately in at least two independent experiments with n ≥ 2 per group. P values determined by one-way ANOVA (a) or two-tailed Mann–Whitney test (d, f–h, j, l, o).
Extended Data Fig. 3 Host-derived IL33 activates pancreatic T cell immunity.
a, Gene set enrichment analysis of bulk RNA-seq from purified CD45+ immune cells from Il33+/+ and Il33−/− PDAC mice. Enrichment plots and enrichment scores are shown for three gene sets comparing expression in Il33−/− to expression in Il33+/+ mice (n = 3 mice per group). FDR, false discovery rate. b, c, Gating of CD8+ T cells (b) and frequencies of various immune cell types (c, left) and CD4+ T cell lineages (c, right) in Il33+/+ and Il33−/− orthotopic PDAC mice. d, Frequency of T central memory (TCM) cells (CD45+ CD3+ CD8+ CD44+ CD62L+) in tumour DLNs and non-tumour draining distant lymphoid organs (inguinal lymph node and spleen) in Il33+/+ and Il33−/− orthotopic PDAC mice. e, Frequency of CD8+ T cells in subcutaneous PDAC tumours. NK, natural killer cells; NKT, natural killer T cells; Treg, regulatory T cells; MDSC, myeloid-derived suppressor cells; DC, dendritic cells. Data were collected 14 days after tumour implantation or at the time points indicated. Median ± s.e.m; horizontal bars show median. n indicates individual mice analysed separately in at least two independent experiments with n ≥ 2 per group. P values determined by one-way ANOVA (d).
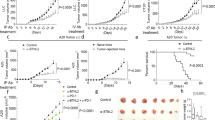
Extended Data Fig. 4 IL33 and ILCs do not directly induce tumour cell death.
a, Tumour weight in Rag2−/− and Rag2−/−γc−/− PDAC mice treated with vehicle or rIL33. b, Representative haematoxylin and eosin-stained sections (left) with histologic tumour cell differentiation status in Il33+/+ and Il33−/− PDAC mice (right). c, Trichrome staining in tumours from Il33+/+ and Il33−/− PDAC mice (n = 3 per group). d, Immunohistochemistry for smooth muscle actin in tumours from Il33+/+ and Il33−/− PDAC mice (n = 3 per group). e, Intratumoral ST2 expression on KPC cells in Il33+/+ and Il33−/− orthotopic PDAC mice. f, ST2 expression on live KPC cells following rIL33 treatment in vitro (DRAQ7 stains dead cells) (n = 3 per group). g, KPC cell number, viability, proliferation (Ki-67), and apoptosis (annexin) following rIL33 treatment in vitro (n = 3–6 per group). Horizontal bars show median. n in a–e indicates individual mice analysed separately in at least two independent experiments with n ≥ 3 per group. n in f, g indicates technical replicates and is representative of at least two independent experiments. P value determined by two-tailed Mann–Whitney test (a).
Extended Data Fig. 5 ILC2s prime antigen-specific CD8+ T cells.
a, Gating and frequency of TILC2s in ILC2-intact (diphtheria toxin (DT)-treated Cd4Cre/+Icos+/+) and ILC2-depleted (DT-treated Cd4Cre/+Icosfl.DTR/+) mice. b, Gating and frequency of OVA-specific CD8+ T cells in spleens from ILC2-intact and ILC2-depleted mice. OVA-specific T cells were detected as SIINFEKL-tetramer+ cells. c, Gating and frequency of TCM cells (CD45+ CD3+ CD8+ CD44+ CD62L+) in tumour, DLNs, and spleens from ILC2-intact and ILC2-depleted mice. d, ST2 expression on CD45+ CD3+ CD8+ T cells after tumour implantation in PDAC mice. Data were collected 14 days after tumour implantation or at the time points indicated. Median ± s.e.m; horizontal bars show median. n indicates individual mice analysed separately in at least two independent experiments with n ≥ 2 per group. P values determined by two-tailed Mann–Whitney test (a–c) and two-way ANOVA with Tukey’s multiple comparison post test (d, indicating comparison of tumour ILCs to all other groups).
Extended Data Fig. 6 Immunophenotyping in rIL33-treated PDAC mice.
a, Tumour establishment of orthotopic and subcutaneous KPC-OVA PDAC tumours in vehicle (veh) and rIL33-treated mice. b, Gating (left) and frequency (right) of IL18R1 expression on tumour ILCs in subcutaneous (SQ) and orthotopic PDAC mice. c, Gating (left) and frequency (right) of splenic ILC2s following rIL33 treatment in orthotopic PDAC mice. d, Gating (left), frequency (middle), and number (right) of TILC2s following rIL33 treatment in subcutaneous PDAC mice. e, Gating (left) and frequency (right) of cytokine and PD-1 expression on tumour CD8+ T cells following rIL33 treatment in orthotopic PDAC mice. f, Frequency of immune cells in vehicle- and rIL33-treated orthotopic PDAC mice. g, Gating strategy for identification of CD103+ DCs. h, Gating (left; tumours) and frequency (right) of ILC2s in tumours and DLNs from wild-type (WT) or ILC2-deficient (Rorafl/flIl7rCre) PDAC mice following rIL33 treatment. i, Gating (left) and frequency (right) of PD-1+ CD8+ T cells in tumours from rIL33-treated wild-type (WT) and Batf3−/− mice. Data were collected 6 (a), 4 (b), 2 (c, e, f), 5 (d), 7 (h), and 3 (i) weeks after tumour implantation. Horizontal bars show median. n indicates individual mice analysed separately in at least two independent experiments with n ≥ 2 per group. P values determined by χ2 test (a), two-tailed Mann–Whitney test (all else).
Extended Data Fig. 7 scRNA-seq of tumour and DLN ILC2s in PDAC mice.
a, Experimental design for in vivo treatment, purification, and single-cell analysis of ILC2s. b, c, Quality metrics. b, Scatter plots showing, for each cell, the relationship between the number of UMIs and the number of genes. c, Violin plots showing the distribution of the number of genes (left), number of UMIs (middle), and percentage of normalized reads from mitochondrial genes (right) in each treatment group (columns), and each tissue (rows). Each dot represents a single cell. For each treatment group and tissue, data represent pooled purified single cells from biological replicates of n = 10 (vehicle), n = 5 (rIL33), and n = 5 (anti-PD-1 + rIL33) PDAC mice.
Extended Data Fig. 8 Activated ILC2s from tumours and DLNs have distinct transcriptional features.
a–d, Single-cell analysis of 1,634 rIL33-activated tumour and DLN ILC2s (experimental design as in Extended Data Fig. 7a). UMAP plots show single cells (dots) in a nonlinear representation of the top 15 principal components. Expression of ILC2 (Gata3, Id2, Rora) and ILC3 (gene, Rorc; protein, RORγT) transcription factors (TFs) (a), ILC2 surface markers (b), and ILC clusters and tissues (tumour and DLN) (c). Expression of the ILC-1 transcription factor Tbx21 (T-bet) was undetectable. d, e, Differentially expressed genes by cluster (d) and tissue (e). f, Distribution of Ccl5 expression from ILC2s in tumours and DLNs; violin plots show distribution with minima, maxima, and circle indicating median. Each dot in a–c represents a single cell. For each treatment group and tissue, data represent pooled purified single cells from biological replicates of n = 5 rIL33-treated PDAC mice. P value in f by two-sided pairwise Wilcoxon rank sum test.
Extended Data Fig. 9 Combined anti-PD-1 and rIL33 treatment induces a unique transcriptional profile in TILC2s.
a, Expression of coinhibitory immune checkpoints in TILC2s in vehicle-treated PDAC mice by scRNA-seq. b, Gating and frequency of PD-1+ ILC2s in vehicle- and rIL33-treated PDAC mice. c, ILC2 frequency in treated PDAC mice. Corresponding tumour volumes, cell number, and scRNA-seq are shown in Fig. 4a–c. d, scRNA-seq of ILC2s from treated PDAC mice. Expression of ILC1 (gene, Tbx21; protein, T-bet), ILC2 (Gata3, Id2, Rora), and ILC3 (gene, Rorc; protein, Rorγt) transcription factors (TFs) in purified tumour and DLN ILC2s. Corresponding UMAP plots by cluster and treatment are depicted in Fig. 4c. e–g, Top differentially expressed genes by treatment and tissue (e), cluster (f), and distribution of expression for select differentially expressed genes by treatment and tissue (g). h, UMAP plots of 3,415 single TILC2s in a nonlinear representation of the top 15 principal components. i, Differentially expressed genes in TILC2s by treatment. Each dot in d, h represents a single cell; in d–i, for each treatment group and tissue, data represent pooled purified single cells from biological replicates of n = 10 (vehicle), n = 5 (rIL33), and n = 5 (anti-PD-1 + rIL33) PDAC mice (number of single cells for tumour: vehicle n = 28, rIL33 n = 752, rIL33 + anti-PD-1 n = 2,635; for DLN: rIL33 n = 882, rIL33 + anti-PD-1 n = 2,725). Violin plots show distribution with minima, maxima, and circle indicating median. Horizontal bars show median. P values by two-tailed Mann–Whitney test (b, c) and two-sided pairwise Wilcoxon rank sum test (g).
Extended Data Fig. 10 Activated TILC2s express PD-1 and co-exist with PD-1+ T cells.
a, b, Orthotopic PDAC mice (C57BL/6 WT, Pdcd1−/−, CD45.1) were treated with 500 ng of carrier-free rIL33 daily for 10 days (experimental designs shown in Fig. 4e, f). Live, CD45+, lineage−, CD90+, CD25+, ST2+ TILC2s were sort-purified to 98% purity at day 10 post-implantation. TILC2s (5 × 105 cells) were immediately transferred to orthotopic PDAC tumour-bearing ILC2-deficient (Rorafl/flIl7rCre/+) CD45.2 mice on days 7 and 14 post-tumour implantation via i.p. injection. Control mice received equivalent volumes of PBS via i.p. injection. a, Representative plots for TILC2 sort-purification (top) and post-sort purity (bottom). b, Representative plots showing PD-1 expression on sort-purified TILC2s from wild-type and CD45.1 mice in the experimental designs outlined in Fig. 4e, f. c, Survival and intratumoral CD8+ T cell frequency of orthotopic KPC 4662 and KPC 52 PDAC tumours; horizontal bars in c show median. d, Frequency of PD-1+ ILC2s (left) and correlation with PD-1+ T cells (right) in human PDACs. e, Linear regression analysis of IL33 and PDCD1 mRNA in bulk tumour transcriptomes from short- and long-term human PDAC survivors (left) and survival association of PD-1+ cells in tumour tissue microarrays of short- and long-term PDAC survivors (right); high and low defined as higher or lower than the median for the cohort. f, Model linking the IL33–TILC2 axis to T cell immunity in PDAC. ©2019, Memorial Sloan Kettering Cancer Center. g, Distribution of expression of costimulatory molecules in untreated TILC2s by scRNA-seq. Experimental design as shown in Extended Data Fig. 7a; data represent pooled purified single cells from biological replicates of n = 10 (vehicle). Data are representative of purity and PD-1 expression on sorted TILC2s in two independent experiments with n ≥ 4 per group (a, b). n and data points denote individual mice and patients analysed separately. P values determined by two-tailed Mann–Whitney test (c), two-sided log rank test (c, e, survival curves) and linear regression (d, e).
Supplementary information
Supplementary Tables
Supplementary Tables: Contains three tables with data on – (1) the clinicopathological characteristics of PDAC patients in the cohort analysed by flow cytometry (Fig. 1a); (2) the clinicopathological characteristics of PDAC patients in transcriptome cohort (Fig. 1c, Extended Data Fig. 1c); and (3) ILC2 single-cell RNA sequencing cellular and transcriptomic metrics (Fig. 4c, Extended Data Figs. 7-9, 10g).
Supplementary Data
Supplementary Data: Source code for immune quantification.
Source data
Rights and permissions
About this article

Cite this article
Moral, J.A., Leung, J., Rojas, L.A. et al. ILC2s amplify PD-1 blockade by activating tissue-specific cancer immunity. Nature 579, 130–135 (2020). https://doi.org/10.1038/s41586-020-2015-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41586-020-2015-4
This article is cited by
-
Cytolytic too: Granzyme B-expressing human ILC2s mediate tumor rejection
Cell Research (2024)
-
A diversity of novel type-2 innate lymphoid cell subpopulations revealed during tumour expansion
Communications Biology (2024)
-
Group 3 innate lymphoid cells in intestinal health and disease
Nature Reviews Gastroenterology & Hepatology (2024)
-
Combination of IL-33 with PD-1 blockade augment mILC2s-mediated anti-tumor immunity
Cancer Immunology, Immunotherapy (2024)
-
Group 2 innate lymphoid cells and their surrounding environment
Inflammation and Regeneration (2023)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
