Article Text

Abstract
Objective Our previous studies have identified CXCL8 as the crucial chemokine responsible for gastric cancer metastasis mediated by loss of RACK1. However, the regulatory effect of CXCL8 on immune surveillance in gastric cancer remains obscure.
Design Flow cytometry analyses were performed to examine major source of CXCL8 and phenotypes of immune cells in fresh tumour tissues from 76 patients with gastric cancer. Real-time PCR was performed to analyse CXCL8 mRNA level in gastric cancer tissues. For immunohistochemical analyses, a total of 420 patients with gastric cancer undergoing curative resection were enrolled. In vitro culture of fresh tumour tissue was performed to evaluate the potential therapeutic effect of blocking CXCL8 pathway in gastric cancer.
Results Increased level of CXCL8 indicates poor clinical outcome and tumour progression in patients with gastric cancer. In gastric cancer tissues, CXCL8 is predominantly secreted by macrophages and colony stimulating factor 2 (CSF-2) facilitates macrophage-derived CXCL8 secretion. High level of CXCL8 is associated with decreased CD8+ T cells infiltration and Ki67+ CD8+ T cells proportion. Moreover, CXCL8 also inhibits CD8+ T cells function by inducing the expression of PD-L1 on macrophages. Finally, we show that a small-molecule CXCR2 inhibitor, reparixin, drives the decreased programmed death-ligand 1 (PD-L1+) macrophages and promotes antitumour immunity. Accordingly, high levels of CXCL8+ macrophages are positively correlated with poor prognosis in patients with gastric cancer.
Conclusions CXCL8 is predominantly secreted by macrophages and contributes to the immunosuppressive microenvironment by inducing PD-L1+ macrophages in gastric cancer. CXCL8 inhibitors may drive antitumour response, providing potential therapeutic effects for patients with gastric cancer.
- gastric cancer
- immunotherapy
- macrophages
Statistics from Altmetric.com
Significance of this study
What is already known on this subject?
Immune checkpoint inhibitors have showed promising efficacy and manageable safety in patients with gastric cancer.
M2 type macrophages infiltration is associated with an unfavourable clinical outcome in gastric cancer.
CXCL8 has been confirmed to promote gastric cancer metastasis by inhibiting RACK1.
What are the new findings?
In gastric cancer tissues, CXCL8 is predominantly secreted by macrophages and CSF-2 facilitates macrophage-derived CXCL8 secretion.
High level of CXCL8 is associated with decreased CD8+ T cells infiltration and Ki67+ CD8+ T cells proportion.
CXCL8 inhibits CD8+ T cells function by inducing the expression of PD-L1 on macrophages.
Blockade of CXCL8 pathway contributes to the decreased PD-L1+ macrophages and inhibits immune evasion.
How might it impact on clinical practice in the foreseeable future?
Our data provide evidence that tumour-associated macrophages-derived CXCL8 determines immune evasion through autonomous PD-L1 expression in gastric cancer. The findings suggest that it may be a promising strategy to block the CXCL8 pathway to increase antitumour immunity in gastric cancer.
Introduction
Gastric cancer is the fifth most common cancer in the world and the second leading cause of cancer death. Nearly 1 million new diagnoses are made every year. More than one-third of the world total occurs in China.1 Radical gastrectomy is the only curative treatment for gastric cancer.2 3 In patients who have undergone adequate complete surgical resection (R0) of locally advanced gastric cancer, adjuvant chemotherapy has been generally accepted to improve 5-year overall survival (OS) by 10%–15%.4–6 Although traditional treatments do bring survival benefits to the patients with gastric cancer, many patients experience relapse soon after postoperative adjuvant therapy indicating that the tumour heterogeneity facilitates the escape from cytotoxic therapies.
The results of KEYNOTE-059 and CheckMate 032 study have opened a brand new chapter in the war against gastric cancer. Immune checkpoint inhibitors showed promising efficacy and manageable safety in patients with gastric cancer, which indicates that the focus of therapy has shifted from the tumour itself to the microenvironment, to mobilise immune cells to recognise and finally kill the cancer cells.
The crosstalk between myeloid cells and adaptive immunity play a key role in the tumour progression, with tumour infiltrating myeloid cells possibly having an important role in immunosuppression. Macrophages are an abundant population of leucocytes in gastric cancer. They could be classified into two extreme forms referred to as ’classical' (or M1) and ’alternative' (or M2) activation. These two states are defined by responses to the cytokine interferon (IFN)-γ and activation of toll-like receptors (TLRs), and to interleukin (IL)-4 and IL-13, respectively. M1 macrophage activation increases their ability to present antigens via major histocompatibility complex (MHC) class II molecules and kill pathogens. M2 activation state is characterised by activation of metabolic pathways that can inhibit adaptive immune responses. Previously, we have showed increased M2 type macrophages infiltration was associated with an unfavourable clinical outcome in gastric cancer. More recently, it has been demonstrated that M1 and M2 profiles are artificial in vitro extremes and tumour-associated macrophage (TAM) phenotypes typically go beyond simple M1 and M2 denominations. Instead, from a functional point of view, macrophages with specific phenotype emerge as an important regulator in the tumour progression. Ablation of specific subpopulations including Tie2+ or metastasis-associated macrophage (MAMs) has been demonstrated to affect specific activities such as angiogenesis or metastatic seeding.7 However, direct evidence supporting macrophages with specific phenotype in the immunosuppression of gastric cancer is lacking.
CXCL8 was initially noted for its role in neutrophil exocytosis and oxidative burst in 1980s.8–10 Now accumulated evidence has showed that elevated levels of CXCL8 led to many malignant biological behaviours of cancer cells, which link to poor clinical outcome.11–15 Our previous study found that CXCL8 induced metastasis in gastric cancer.16 However, the impact of CXCL8 on the tumour immunological microenvironment in gastric cancer is poorly understood.
Here, we showed that CXCL8, which correlated with OS in patients with gastric cancer, was predominantly expressed by macrophages. Furthermore, we demonstrated that CSF-2 promoted macrophage-derived CXCL8 secretion and CXCL8 subsequently induced reduced CD8+ T cell infiltration and increased PD-L1 expression on macrophages, which inhibited CD8+ T cells activity. Our research provided direct evidence for the important role of CXCL8 in the immunosuppression of gastric cancer.
Materials and methods
Patients and follow-up
Two independent cohorts of patients with gastric cancer were enrolled in the study. One cohort including 420 patients with gastric cancer who underwent radical gastrectomy between 2007 and 2008 was obtained from Zhongshan Hospital, Fudan University. At the begining, the study recruited 496 patients. However, 468 of the 496 patients have comprehensive information about chemotherapy, clinicopathological data, and survival outcomes for complete analysis. In this study, we utilised the tissue microarrays (TMAs) constructed by Shanghai Outdo Biotech Co, Itd, and 40 dots of the 468 patients were lost after immunohistochemistry. Additionally, we excluded 8 patients of TNM stage IV. Consequently, we included 420 patients in this study. All patients were followed up until April 2014. OS was calculated from the date of gastrectomy to the date of death or last follow-up. None of the patients received preoperative chemotherapy. Written consent was obtained from each patient. The clinical stages of tumours were determined according to the tumour, node, metastases (TNM) classification system of the International Union Against Cancer (seventh edition). The other cohort (TCGA) comprising 384 patients with gastric cancer was obtained from TCGA database.
Immunohistochemistry and immunofluorescence
Tissue microarray construction and the immunohistochemistry (IHC) protocol have been described previously.17 For immunohistochemical staining, sections were exposed to the primary antibody at 4°C overnight. The antibodies against CD68, CSF-2, CXCL8, CD8, PD-L1 were obtained from Abcam. Slides were then treated with polyperoxidase-conjugated IgG (OriGene). Staining was performed with diaminobenzidine solution (Biocare Medical) under a microscope and counterstained with haematoxylin. The primary antibody was omitted for the negative controls. Two pathologists, blinded to patient characteristics, evaluated the staining of each specimen. In case of disagreement, the slides were reviewed and a consensus was reached by the two observers.
For immunofluorescence staining, the sections were incubated with the primary antibodies overnight at 4°C. Then samples were then incubated with species-appropriate rabbit secondary antibodies coupled to AlexaFluor dyes (488, 594, Invitrogen) for 1 hour at room temperature. VECTASHIELD with Hoechst (Vector Laboratories) was used to mount cover slips. The slides were analysed using Leica DMi8 microsystems.
Flow cytometry
For fresh tumour tissues, single cells were isolated by using collagenase IV and then stained with appropriate monoclonal antibodies (mAbs) for 30 min at 4°C. When necessary, staining of intracellular protein was performed by use of the Fixation/Permeabilization Solution Kit (BD Biosciences) according to the manufacturer’s instructions. Fluorochrome-conjugated CD45, CD8, CD68, CD4, CD66b, CD326, Annexin V, GZMB, Ki67 and PD-L1 mAbs were purchased from BD Pharmingen. Stained cells were washed, re-suspended in phosphate-buffered saline/0.1% bovine serum albumin plus azide. Flow cytometry was performed using an BDcelesta and analysed by FlowJo software (Tree Star).
ELISA
CXCL8 were measured by ELISA in supernatants of macrophages induced from peripheral blood mononuclear cell (PBMC) with or without recombinant human proteins CSF-3, CSF-2, CXCL1 (R&D Systems) and tumour tissue supernatant with IgG or neutralising anti-CSF-2. ELISA kit used was the human CXCL8 kits obtained from R&D Systems.
In vitro tumour inhibition assay
Fresh gastric cancer tissues were washed two times with RPMI-1640 medium containing 1% fetal bovine serum before being cut into small pieces. The specimens were then collected in RPMI-1640 medium containing 1 mg/mL collagenase IV and mechanically dissociated using the gentle MACS Dissociator (Miltenyi Biotec) for 3 min. Dissociated cell suspensions were further incubated for 2 hour at 37°C under continuous rotation. The cell suspensions were then filtered through a 40 µm cell strainer (BD Labware) and collected. In another CD8+ T cells-deprived system, CD8+ T cells were depleted from single cell suspensions by negative selection using CD8 MicroBeads (Life Technologies). Single cell suspensions were cultured in assay medium (RPMI-1640) in 6-well plates in a total volume of 2 mL, supplemented with 100 IU/mL rhIL-2 under 37°C and 5% CO2. Then the cells were cultured with reparixin (8 µg/mL) or vehicle for 12 hours. After overnight culture, the cells were subjected to phenotypic analysis by flow cytometry as above.
Real-time PCR with reverse transcription
Total RNA from clinical specimens was extracted using TRIzol reagent (Invitrogen) according to the manufacturer’s protocol. Real-time PCR was performed on the Applied Biosystems 7300 Real-Time PCR system using SYBR Green dye (Applied Biosystems) as described by the manufacture. The primers for CXCL8 used are: 5': CACTGCGCCAACACAGAAAT and 3': GCCCTCTTCAAAAACTTCTCCAC. All determinations were performed in triplicate and in at least three independent experiments.
Gene set enrichment analysis
Gene set enrichment analysis (GSEA) performed by the Molecular Signature Database (MSigDB) was used to identify the pathways that were significantly enriched in CXCL8 high tumour samples. If a gene set had a positive enrichment score, the majority of its members had higher expression accompanied with higher risk score, and the set was termed ‘enriched’.
Differential expression analysis
Differential expression analysis of genomic data was performed by Limma moderated t-test. Significantly upregulated and downregulated genes between CXCL8high and CXCL8low tumour samples were defined as expressed genes with false discovery rate (FDR)-adjusted p value <0.05 and fold change ≥2.
Statistical analysis
Results are expressed as mean±SD. Student’s t-test or Wilcoxon rank-sum test was used for continuous variables. Analysis of the correlation was made by Spearman’s correlation. Kaplan-Meier method was used to determine OS. Log-rank test was used to compare survival between two groups. A two-tailed p value <0.05 was considered statistically significant.
Results
CXCL8 is predominantly secreted by macrophages and correlates with tumour progression and poor prognosis in gastric cancer
To elucidate the prognostic value between CXCL8 and clinical outcomes, we performed Kaplan-Meier analysis according to the data from Zhongshan Hospital. We observed high CXCL8 protein level in patients with gastric cancer was associated with short OS (figure 1A). Furthermore, CXCL8 expression was positively correlated with TNM stage (figure 1B), indicating that CXCL8 may promote tumour progression. CXCL8 may arise from many cells, but the main source of it is still unknown. Flow cytometric analysis demonstrated that CD45+ cells from gastric cancer tissues, but not tumour cells, strongly expressed CXCL8 (figure 1C). This further raises the question that which subpopulation plays a dominant role in producing CXCL8. We found that CXCL8 was predominantly secreted by macrophages based on results of fluorescence-activated cell sorting (FACS) analysis (figure 1D-E). In keeping with this finding, immunofluorescence analysis of CD68 and CXCL8 expression in human gastric cancer tissues also revealed that the majority of CXCL8 was expressed by macrophages (figure 1F). Multivariate analysis revealed that besides tumour invasion and lymph node metastasis, CXCL8 expression was an independent risk factor for OS (figure 1G). Taken together, these results above suggest that CXCL8, dominantly originating in macrophages in gastric cancer, is associated with poor prognosis and tumour progression.

CXCL8 is predominantly secreted by macrophages and correlates with tumour progression and poor prognosis in gastric cancer. (A) Kaplan-Meier plot of cases of gastric cancer based on CXCL8 expression by immunohistochemistry (IHC) in Zhongshan cohort. (B) Relative portion of CXCL8 expression according to tumour, node, metastases (TNM) stage. High CXCL8 expression portion was elevated as the TNM stage progressed. *P<0.05, **p<0.01, for groups connected by horizontal lines. (C) Representative flow cytometry analysis of the CXCL8+ cells from fresh human gastric cancer samples. (D–E) Representative figures and statistics analysis of flow cytometry of CXCL8 producing subpopulation infiltration into gastric cancer (n=6). Bar plots show mean±SD ***P<0.001, for groups connected by horizontal lines. (F) Human gastric cancer specimens stained for CXCL8 (red) and CD68 (green). Nuclei counterstained blue with Hoechst. The lower right panel is merged by the aforementioned three images. (G) Cox multivariate analysis identified the independent prognostic factors for overall survival in patients with gastric cancer.
CSF-2 facilitates macrophage-derived CXCL8 secretion
Having identified macrophages as the dominant source of CXCL8 in gastric cancer, we sought to define the molecular mechanism underlying the strong CXCL8 secretion in macrophages. We used GSEA to identify pathways that were activated in CXCL8 high tumours as compared with CXCL8 low tumours, aiming to identify potential regulators for CXCL8 secretion and identified cytokine pathways and cytokine-cytokine receptor interaction as two of the most significantly upregulated pathways (figure 2A). Thus, we hypothesised that cytokines induced macrophage-derived CXCL8. To identify the potential cytokine regulating macrophage-derived CXCL8, we performed clustering analysis based on CXCL8 expression from TCGA database. Comparisons revealed various cytokine genes that were expressed differentially by CXCL8 high tumours relative to CXCL8 low tumours. The top 10 upregulated cytokine genes in CXCL8 high tumours were shown in the heatmap (figure 2B). To identify the exact cytokine facilitating macrophages to secrete CXCL8. We selected the top three upregulated cytokines in CXCL8 high tumours (CSF-3, CSF-2 and CXCL1) to stimulate macrophages induced from PBMCs and detected the concentration of CXCL8. We observed that CXCL8 concentration was significantly increased by the addition of recombinant human CSF-2 in monoculture (figure 2C). To further ensure CSF-2 could induce increased secretion of CXCL8, we added anti-CSF-2 to macrophages induced from PBMC with tumour tissue supernatant. As expected, compared with IgG, anti-CSF-2 neutralising antibody significantly decreased CXCL8 concentration (figure 2D). Moreover, significant correlations were found between the levels of CXCL8 and CSF-2 expression both in protein level from IHC and mRNA level from TCGA database (figure 2E and F).

CSF-2 facilitates macrophage-derived CXCL8 secretion. (A) Gene set enrichment analysis (GSEA) revealed an enrichment of cytokine pathway and cytokine-cytokine receptor interaction involved in CXCL8 high tumours. NES, non-enrichment score. (B) Heatmap of the top 10 upregulated cytokines genes overexpressed in CXCL8 high tumours. (C) Statistical analysis of CXCL8 concentrations of macrophages induced from PBMCs after stimulation with recombinant human CSF-3, CSF-2, CXCL1. ***P<0.001, for groups connected by horizontal lines (n=5). (D) Statistical analysis of CXCL8 concentrations of tumour tissue supernatant with anti-CSF-2 or IgG as determined by ELISA. *P<0.05, for groups connected by horizontal lines (n=5). (E–F) Correlations assessed by Spearman’s correlation between mRNA (TCGA database) and protein (immunohistochemistry (IHC)) level of CSF-2 and CXCL8.
CXCL8 contributes to immunosuppressive microenvironment
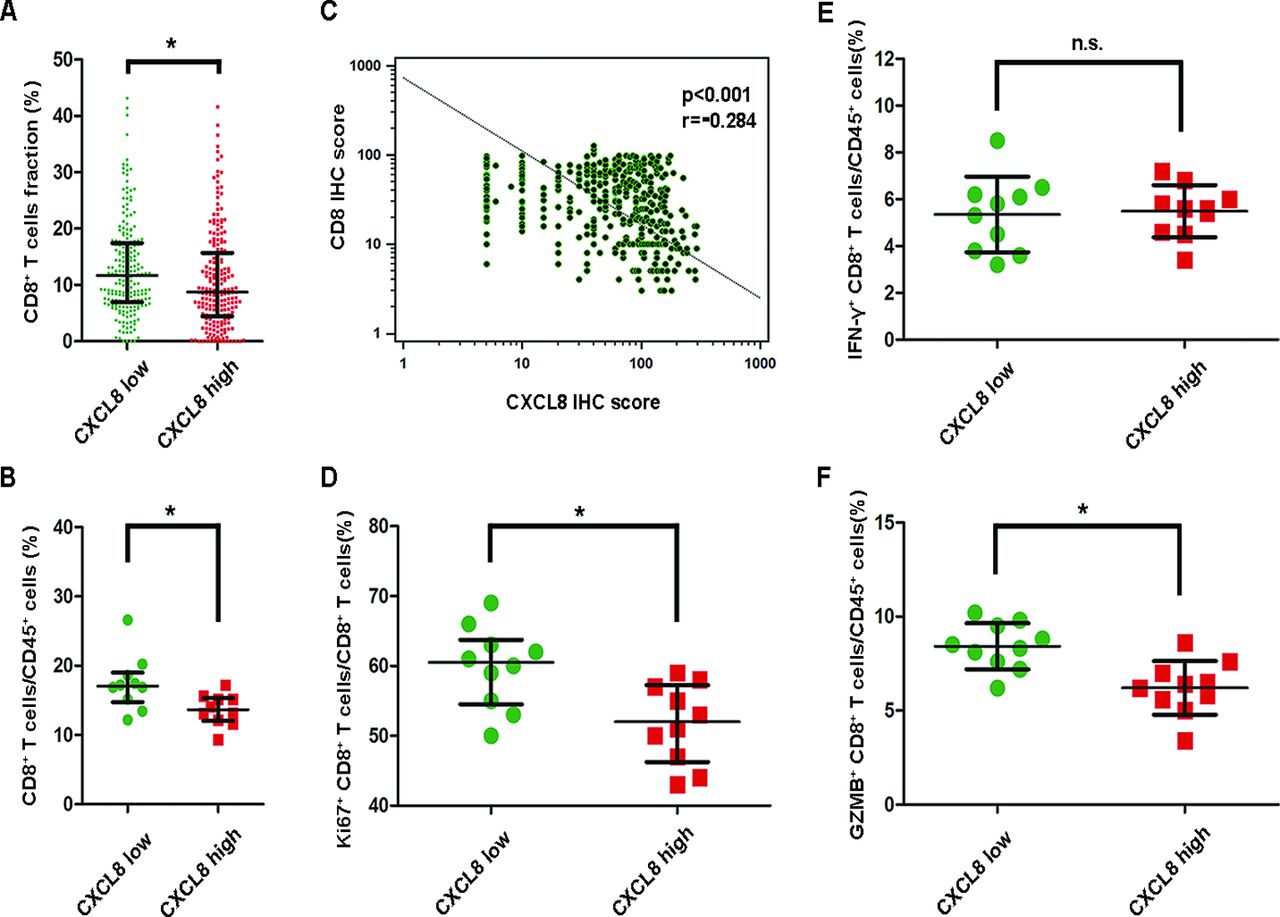
To investigate the influence of CXCL8 on immunosuppression, we analysed CD8+ T cells infiltration in gastric cancer. The consequence of CIBERSORT revealed that patients with high CXCL8 mRNA levels showed a low CD8+ T cells percentage (figure 3A). This is further supported by results of FACS (figure 3B) and IHC (figure 3C) that CXCL8 high tumours were infiltrated with significantly decreased CD8+ T cells. Furthermore, the Ki67+ CD8+ T cell proportion was also reduced in CXCL8 high tumours (figure 3D). Thus, our next objective was to determine the role of CXCL8 in CD8+ T cell function. Interestingly, we found that decreased GZMB+ CD8+ T cells but not IFN-γ+ CD8+ T cells percentage was associated with high CXCL8 levels (figure 3E and F). Collectively, these findings suggest that CXCL8 high tumours contributes to immunosuppressive microenvironment.

CXCL8 contributes to immunosuppressive microenvironment. (A) Frequency of CD8+ T cell populations calculated using CIBERSORT according to TCGA data, with difference in populations computed using unpaired t-test. (B) Tumour cell suspensions from patients with gastric cancer (n=20) were surface stained for CD8 and CD45. CXCL8 high tumours were infiltrated with significantly decreased CD8+ T cells. (C) Correlations assessed by Spearman’s correlation between densities of CD8+ T cells and CXCL8 expression in patients with gastric cancer based on the results of immunohistochemistry (IHC). (D) Ki67+ CD8+ T cells frequencies of CD8+ T cells in gastric cancer tissue samples from patients with CXCL8 high tumours (n=10) or CXCL8 low (n=10) were compared (two-tailed student’s t-test). (E–F) Interferon (IFN)-γ+ CD8+ T cells and GZMB+ CD8+ T cells frequencies of CD45+ leucocytes in gastric cancer tissue samples from patients with CXCL8 high tumours (n=10) or CXCL8 low (n=10) were compared (two-tailed student’s t-test). Bar plots show mean±SD in panels A, B, D, E, F. *P<0.05, **p<0.01, ***p<0.001, n.s. not significant for groups connected by horizontal lines.
CXCL8 promotes immunosuppression by inducing PD-L1+ macrophages
To test how CXCL8 suppresses CD8+ T cells function, we analysed the differentially expressed gene based on TCGA database. We observed that PD-L1 was overexpressed in CXCL8 high tumours (figure 4A). This was further supported by results of FACS analysis of tumour suspension, which showed CXCL8 expression was positively associated with PD-L1 level (figure 4B-C). In addition, FACS analysis also showed that macrophages were the main source of PD-L1 in gastric cancer (figure 4D-4E). In accordance with this phenomenon, a significant correlation between CD68 and PD-L1 was observed in protein and mRNA level (figure 4F-4G). This raised a question that whether CXCL8 was associated with PD-L1 expression on macrophages. Interestingly, using FACS, we confirmed that CXCL8 high tumours were infiltrated with significantly increased PD-L1+ macrophages (figure 4H-4I). These results suggest that CXCL8 modulates immunosuppressive microenvironment by inducing PD-L1+ macrophages.

CXCL8 promotes immunosuppression by inducing PD-L1+ macrophages. (A) Volcano plot shows differentially expressed genes between CXCL8 high tumours relative to CXCL8 low tumours. mRNA levels with a p<0.05 and fold change ≥2 or ≤0.5 were deemed differentially expressed. (B–C) Representative FACS figures and statistics analysis of PD-L1+ cells frequencies in gastric cancer tissue samples from patients with CXCL8 high tumours (n=10) or CXCL8 low tumours (n=10). Colour histograms represent staining of PD-L1; grey. CXCL8 mRNA expression were compared (two-tailed student’s t-test). (D–E) Representative figures and quantification by flow cytometry of PD-L1 producing subpopulation infiltration into gastric cancer tissues (n=10). (F–G) Correlations assessed by Pearson’s correlation analysis and linear regression analysis between densities of macrophages and PD-L1 expression in patients with gastric cancer based on the results of immunohistochemistry (IHC). (H–I) Representative figures and quantification by flow cytometry of PD-L1+ macrophages frequencies of total macrophages in gastric cancer tissue samples from patients with CXCL8 high (n=10) or CXCL8 low (n=10). Colour histograms represent staining of PD-L1; grey. CXCL8 expression were compared (two-tailed student’s t-test). Bar plots show mean±SD in panels C, E, I. *P<0.05, **p<0.01, ***p<0.001, for groups connected by horizontal lines. FMO, fluorescence minus one.
Blockade of CXCL8 pathway increases antitumour immunity
To further ensure that CXCL8 could induce PD-L1 on macrophages, we added reparixin (a CXCR1/2 small molecular inhibitor) to gastric cancer suspension and detected PD-L1+ macrophages percentage. As expected, PD-L1+ macrophages were significantly reduced if the tumour suspension was treated with reparixin (figure 5A). Consistent with the result, we observed decreased total PD-L1+ cells in reparixin group (figure 5B), suggesting reparixin may suppress antitumour immunity. To demonstrate the hypothesis, we detected the GZMB+ CD8+ T cells, Ki67+ CD8+ T cells proportion and Annexin V+ and Ki67+ epithelial cells frequencies in gastric cancer tissue samples in the tumour suspension treated with reparixin. Interestingly, we observed increased GZMB+ CD8+ T cells and Ki67+ CD8+ T cells proportion in reparixin group (figure 5C and D). Meanwhile, reparixin caused decreased Ki67+ (figure 5E) and increased Annexin V+ (figure 5F) epithelial cells frequencies. As shown in figure 5F, this increased antitumour effect of reparixin was abolished when CD8+ T cells were depleted from single cell suspension. Moreover, single cells suspension with CD8+ T cells depletion alone did not show a significantly decreased tumour cell apoptosis compared with those without depleting CD8+ T cells. These findings suggest that blocking CXCL8 pathway may promote antitumour immunity.

Blockade of CXCL8 pathway increases antitumour immunity. (A–B) PD-L1+ cells frequencies of CD45+ leucocytes and PD-L1+ macrophages of max cells in gastric cancer tissue samples from patients after treated reparixin or dimethyl sulfoxide (DMSO) (n=8). (C–D) GZMB+ CD8+ T cells frequencies of CD45+ leucocytes and Ki67+ CD8+ T cells frequencies of CD8+ T cells in gastric cancer tissue samples from patients after treated reparixin or DMSO (n=8). (E) Ki67+ epithelial cells frequencies of epithelial cells in gastric cancer tissue samples from patients after treated reparixin or DMSO (n=8). (F) Annexin V+ epithelial cells frequencies of epithelial cells in gastric cancer tissue samples from patients in absence or presence of CD8+ T cells after treated reparixin or DMSO (n=8). Bar plots show mean±SD in panels A–F. *P<0.05, **p<0.01, for groups connected by horizontal lines. n.s., not significant.
CXCL8-PD-L1 axis indicates poor prognosis in gastric cancer
Considering the fact that CXCL8 may suppress antitumour immunity by inducing PD-L1, we analysed potential prognostic significance of CXCL8+ macrophages and PD-L1 in gastric cancer. Gastric cancer samples with high PD-L1 expression or infiltrated with increased CXCL8+ macrophages displayed poor prognosis (figure 6A and B). These findings prompted us to investigate the correlation between CXCL8 and PD-L1. In IHC, CXCL8 expression showed significant positive correlation with PD-L1 within tumours (figure 6C). In accordance with the result, CXCL8 transcripts correlated positively with PD-L1 based on TCGA database (figure 6D).
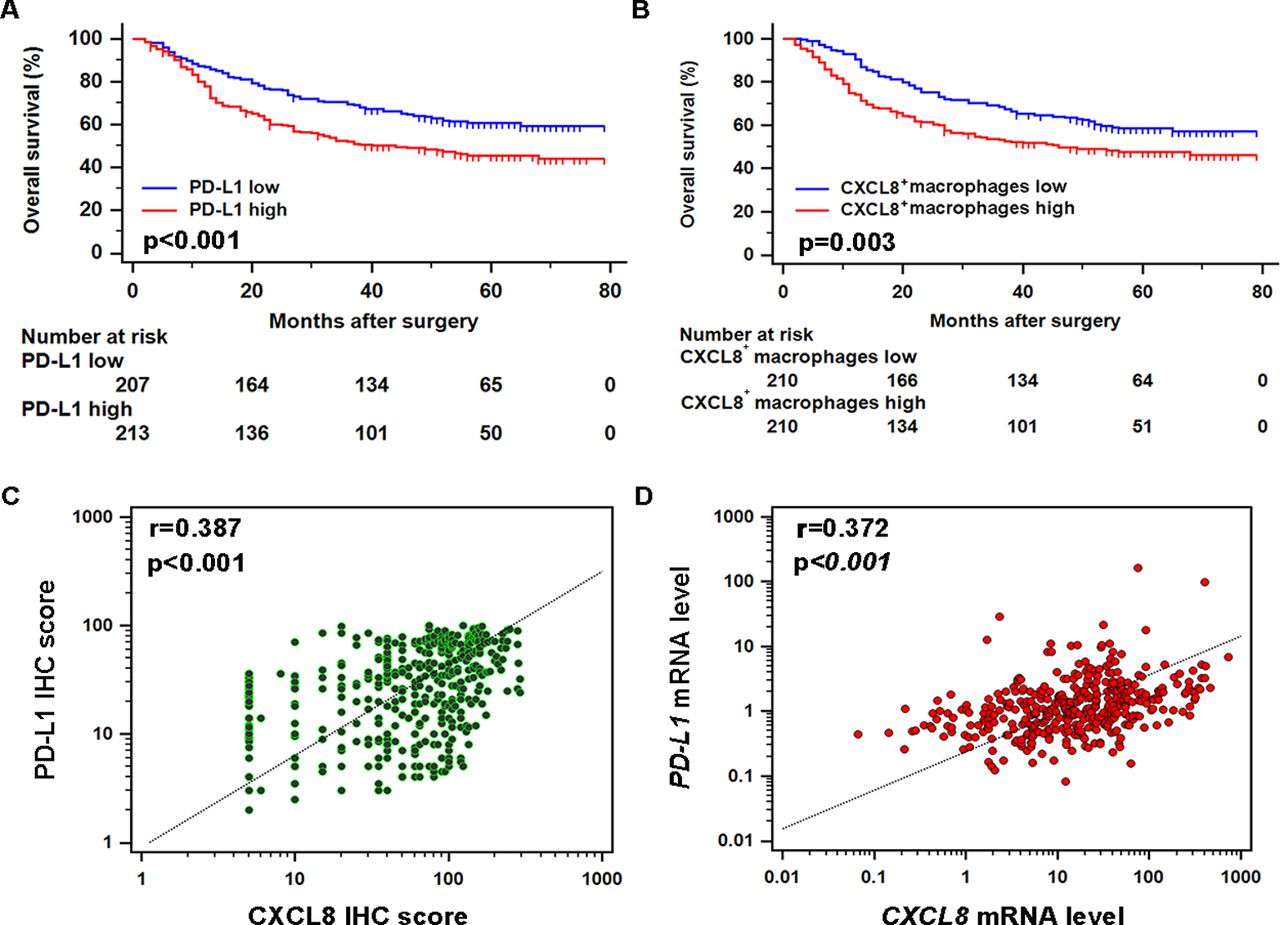
CXCL8-PD-L1 axis indicates poor prognosis in gastric cancer. (A–B) Kaplan-Meier plot of cases of gastric cancer based on PD-L1 expression and CXCL8+ macrophage infiltration. (C–D) Correlations assessed by Spearman’s correlation between mRNA (TCGA database) and protein (immunohistochemistry (IHC)) level of CXCL8 and PD-L1 expression.
Discussion
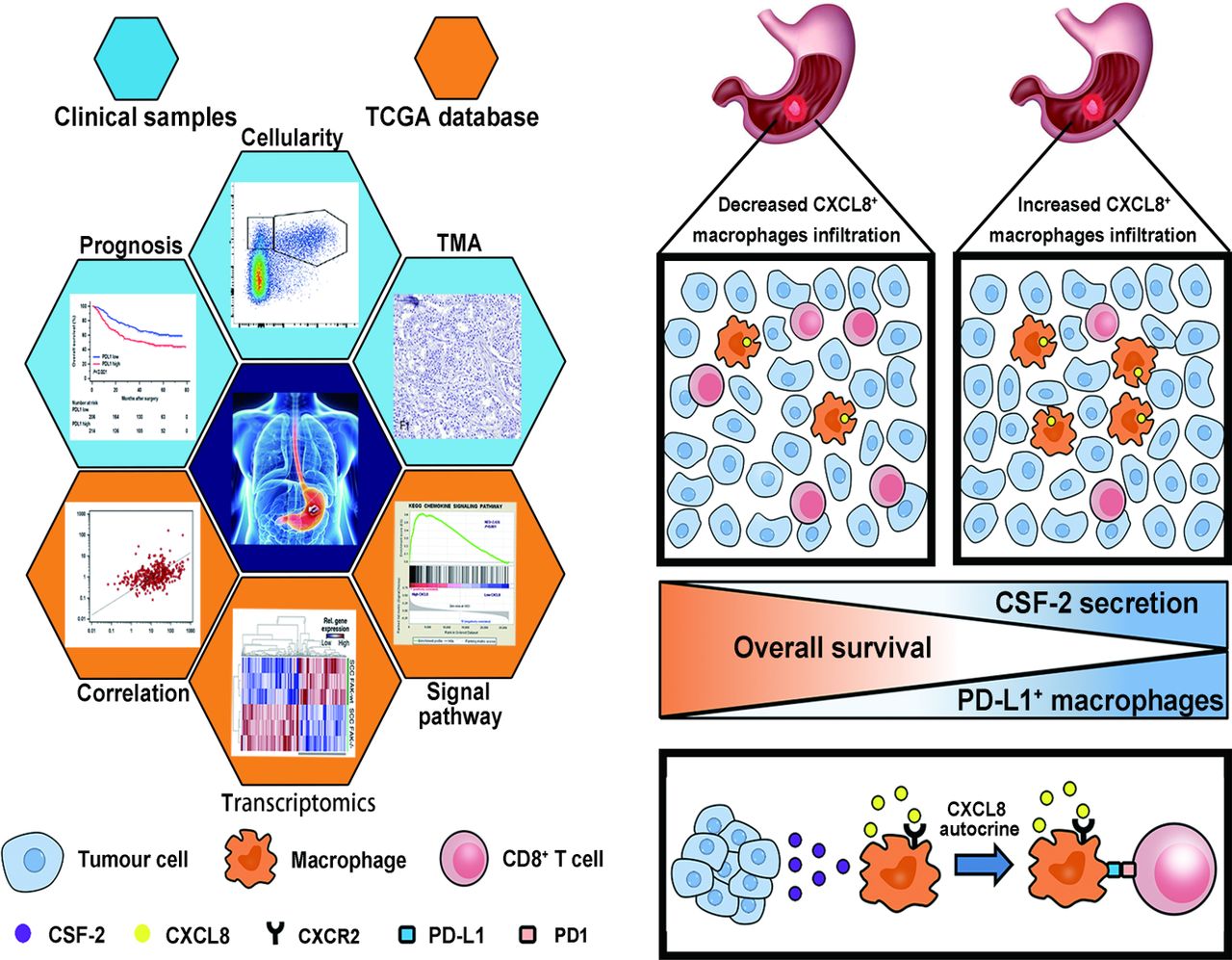
Although substantial evidence has showed a direct relationship between CXCL8 and a poor clinical outcome in gastric cancer. It has been reported that CXCL8 can be produced by macrophages, epithelial cells, airway smooth muscle cells and endothelial cells, but little is known about the major source of CXCL8 in gastric cancer. In this study, we identified macrophages as the dominant origin. Further investigation suggested that CSF-2 facilitated macrophage-derived CXCL8 secretion. CXCL8 is typically known to promote angiogenesis in many solid malignancies. However, the role of CXCL8 in tumour immunosuppression is to be elucidated. In this study, we demonstrated that CXCL8 was associated with decreased CD8+ T cells infiltration and inhibits CD8+ T cell function by inducing PD-L1 on macrophages in gastric cancer. These findings implied that CXCL8 producing macrophages contribute to tumour progression by inducing immunosuppressive microenvironment. This conclusion is supported by our observations that CXCL8+ macrophages infiltration in gastric cancer was negatively associated with overall survival (figure 7).
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Graphical summary of this study. Schematics depicting the materials and methods used in the research and the role of CXCL8+ macrophages in mediating immune escape, which leads to poor prognosis in gastric cancer. TMA, tissue microarray.
TAMs contribute to the suppression of effective adaptive immunity. Macrophages produce IL-10 and transforming growth factor-β to promote the activity of regulatory T (Treg) cells.18 19 Moreover, TAMs result in T cell dysfunction by express a variety of immunoregulatory proteins such as arginase 1,20 21 indoleamine 2,3dioxygenase,22 PD-L1.23 Indeed, evidence indicates that the relative importance of distinct pathways of macrophage differentiation varies between different tumours, resulting in heterogeneous TAM phenotypes and functions. Considering the heterogeneity of TAMs, we infered that the above functions may be performed by different subpopulation of TAMs. On the basis of the fact that macrophages were the dominant source of CXCL8, we wondered whether CXCL8+ macrophages contributed to immunosuppression in gastric cancer. Accordingly, our research provided evidence that CXCL8+ macrophages induced a suppressive immunological environment by inducing PD-L1.
Although previous studies have shown that CXCL8 and its receptors CXCR1/2 are physiologically responsible for the activation and trafficking of granulocytes and neutrophils to the site of inflammation.24 We are the first to report that CXCL8 is predominantly secreted by macrophages in gastric cancer. In accordance, CXCL8 is one of the most abundant cytokines in TAMs in papillary thyroid carcinoma, mediating TAM-dependent tumour invasion and metastasis.25 Interestingly, besides macrophages, we have found neutrophils and CD4+ T cells also express CXCL8, which is consistent with the previous study showing neutrophils and CD4+ T cells could express CXCL8 in endometriosis patients.26 Furthermore, a recent study has proved that Tregs produce high amounts of CXCL8.27 Of course, whether the CXCL8 expressing CD4+ T cells in gastric cancer tissues are Tregs is to be demonstrated. It has been demonstrated that increased CXCL8 secretion is associated with malignant biological functions such as angiogenesis, migration and resistance to anoikis of tumour cells. Moreover, it also degrades extracellular matrix by activating matrix metalloproteinase, which promotes metastasis.28 However, very little is currently known about the role of CXCL8 in immunosuppression. Consistent with our results, Highfill et al 29 found that inhibiting CXCL8-CXCR2 axis reduced MDSC infiltration and inhibited T -cell activation. To our knowledge, this is the first demonstration that CXCL8 induces PD-L1 on macrophages.
CSF-2 promotes macrophage survival and polarisation and induces TAM to differentiate towards M1-like phenotype.18 However, although two extreme forms of polarisation are easy to understand, heterogeneous TAM phenotypes and functions in the tumour microenvironment indicates such definitions are possibly too limiting. Furthermore, Qian showed that macrophages had a mixed phenotype expressing both M1 and M2 markers via large-scale transcriptome analysis.30–32 Our previous study demonstrated that high level of CSF-2 expression predicts poor prognosis in gastric cancer, while Van Overmeire et al 33 showed CSF-2R signalling played a significant role in maintaining the MHC-IIhigh phenotype, whereas the represent study showed CSF-2 increased macrophage-derived CXCL8 secretion. The difference may be methodological. The techniques and specimens used to induce macrophages were different. Overmeire et al used CSF-2 to stimulate monocytes from model mice directly, whereas we induced PBMCs from patients with gastric cancer to differentiate into macrophages by CSF-1 and then stimulated them with CSF-2 in order to imitate the microenvironment in gastric cancer.
The present study clearly has limitations that must be acknowledged. The major source of CSF-2 is still unknown. The concentration of CSF-2 and CXCL8 is positively correlated, the mechanism of CSF-2 promoting CXCL8 secretion is to be explored. Last but not least, whether CXCL8 regulates PD-L1 expression directly or not merit further investigation.
References
Footnotes
CL, HH, HL and RL contributed equally.
Contributors CL, HH, HL and RL for acquisition of data, analysis and interpretation of data, statistical analysis and drafting of the manuscript; YC, YQ, QJ, LC, PZ, HZ and HL for technical and material support; WZ, YS and JX for study concept and design, analysis and interpretation of data, drafting of the manuscript, obtained funding and study supervision. All authors read and approved the final manuscript.
Funding This study was funded by grants from National Natural Science Foundation of China (31470794, 81471621, 81472227, 81501999, 81671628, 81672324, 31770851, 81871306, 81871930) and Shanghai Sailing Program (17YF1402200, 18YF1404600).
Competing interests None declared.
Patient consent Obtained.
Ethics approval The Clinical Research Ethics Committee of Zhongshan Hospital approved the ethical use of human subjects for this study.
Provenance and peer review Not commissioned; externally peer reviewed.