Article Text

Abstract
BACKGROUND AND AIMS The Tage4 gene (tumour associated glycoprotein E4) is overexpressed in rat colon tumours and Min mouse intestinal adenomas. The rat Tage4 protein has approximately 40% identity with human CD155, a member of the immunoglobulin superfamily coding for a transmembrane protein capable of serving as an entry receptor for poliovirus, porcine pseudorabies virus, and bovine herpesvirus 1. Analysis of the ratTage4 gene has revealed structural and functional similarities with the human CD155gene. We therefore investigated expression of theCD155 gene in human colorectal carcinomas.
METHODS Overall CD155 expression was assessed by semiquantitative reverse transcription-polymerase chain reaction (RT-PCR) and immunohistochemical analysis using tissue specimens from patients with colorectal adenomas and adenocarcinomas. We also used a qualitative RT-PCR assay to determine relative expression of different splicing variants in each sample.
RESULTS mRNA levels of CD155 were increased in six of six colorectal cancer tissues compared with the tumour free colon mucosa. Immunohistochemical analysis revealed an increased level of CD155 protein in 12 of 12 samples. The qualitative RT-PCR assay revealed that relative expression of the different CD155 variant transcripts was similar in the different normal and cancer samples tested, indicating that this overexpression is not associated with a particular mRNA variant generated by alternative splicing of the CD155 gene.
CONCLUSION We have shown for the first time that the CD155 gene is overexpressed in colorectal carcinoma and that this overexpression begins at an early stage in tumorigenesis and continues to late stages.
- CD155
- colorectal cancer
- immunoglobulin superfamily
- poliovirus receptor
Abbreviations used in this paper
- CRC
- colorectal carcinoma
- RT-PCR
- reverse transcription-polymerase chain reaction
- CAM
- cellular adhesion molecule
- CEA
- carcinoembryonic antigen
- Ig
- immunoglobulin
Statistics from Altmetric.com
As our understanding of the biology of colorectal cancer progresses, new knowledge about tumorigenesis and tumour biology can be used to diagnose, treat, and prevent this type of cancer. In an initial study of an animal model for immunotherapy of colon tumours, monoclonal antibodies directed against rat colon carcinoma cells were raised.1 One of these monoclonal antibodies (E4) reacts strongly with the primary tumours and lung metastases, as determined by immunohistochemistry. The epithelia of the gastrointestinal tract and of the bronchi are only slightly stained and no staining of other normal rat tissues is observed. Initial experiments have indicated that the antigen recognised by E4 is a transmembrane glycoprotein.2 ,3 The correspondingTage4 gene (tumour associated glycoprotein E4) has since been found to be overexpressed in chemically induced rat colon tumours4 and in Min mouse intestinal adenomas.5 In contrast, a barely detectable level was found in the normal adult rat and mouse tissues tested. TheTage4 cDNA has been isolated.6Sequence analysis using ALIGNp software revealed 40.9% identity between the Tage4 protein and the human CD1557 a member of the immunoglobulin (Ig) superfamily coding for a transmembrane protein capable of serving as an entry receptor for poliovirus, porcine pseudorabies virus, and bovine herpesvirus 1. TheTage4 gene has been mapped to rat chromosome 1q228 and mouse 7A2-B1,9 regions that are homologous to the long arm of human chromosome 19 where theCD155 gene is located.10 To gain insight into the molecular mechanisms involved in its overexpression in cancer cells, we isolated and characterised the ratTage4 gene.11 This gene covers ∼15 kb and is composed of eight exons and seven introns. The humanCD155 gene also contains seven introns in identical positions and phases. In addition, the protein encoded by theTage4 gene was tested for ability to mediate entry of several viruses. The results indicated that theTage4 gene is probably orthologous to the gene for CD155. We therefore decided to analyse expression of theCD155 gene in human colorectal carcinoma (CRC). This was performed by reverse transcription-polymerase chain reaction (RT-PCR) analysis and by immunohistochemistry. We also analysed relative expression of the different variants of theCD155 gene by a qualitative RT-PCR assay.
Materials and methods
TISSUE SAMPLES
Tissue samples were obtained from 18 patients (10 men and eight women; mean age 66 years, range 45–90) undergoing surgical resections for primary CRC diagnosed at the Department of Surgery, Nantes University Hospital, France. Adenomas were collected from four patients: two villous adenoma (both high grade dysplasia) and two tubular adenomas (one low grade and one high grade dysplasia). Sporadic tumours were collected from 14 patients and classified according to Astler and Coller: stage B (n=4), stage C (n=7), and stage D (n=3). The tumours were localised in the right colon (n=5), transverse colon (n=2), left colon (n=3), sigmoid (n=2), and rectum (n=2). Thirteen tumours presented a moderate differentiation, and one was highly differentiated. From each patient tumour tissue and adjacent normal mucosa (about 10 cm from the tumour) were excised and collected. Samples were immediately processed for isolation of epithelial cells and fragments were frozen for immunohistochemistry and kept at −70°C until analysis.
NORMAL COLONIC EPITHELIAL CELLS AND CANCER CELL LINES
Normal epithelial cells were isolated from surgical specimens using a non-enzymatic dissociation technique based on the use of the calcium chelating agent EDTA, as previously described.12
HCT8R, SW620, and SW1116 colon carcinoma cell lines were grown in RPMI-1640 containing l-glutamine and sodium bicarbonate (Sigma, Saint Quentin Fallavier, France) supplemented with 10% fetal calf serum, penicillin (100 units/ml), and streptomycin (0.1 mg/ml). Cells were maintained at 37°C in 5% carbon dioxide and passaged twice a week.
IMMUNOHISTOCHEMISTRY
Cryostat sections (5 μm) were fixed in acetone and stored at −20°C. Before use, slides were rehydrated with phosphate buffered saline (150 mM NaCl, 10 mM phosphate buffer, pH 7.4). The slides were then incubated with normal goat serum for 10 minutes. Mouse monoclonal antibody D171 (Neomarkers, Union City, California, USA) directed against human CD15513 was used as a primary antibody at a concentration of 2.5 μg/ml in antibody diluent containing background reducing components (Dako, Trappes, France). Alkaline phosphatase goat antimouse Ig (Sigma) was used as a secondary antibody (dilution 1:50). Following two washes with phosphate buffered saline and one with Tris buffered saline (150 mM NaCl, 10 mM Tris Cl, pH 7.5), staining was developed using the Vector Red alkaline phosphatase substrate (Vector, Burlingame, California, USA). After staining, tissues were counterstained with Papanicolaou. For control of specific staining an irrelevant monoclonal mouse IgG antibody was used instead of anti-CD155.
RNA PREPARATION AND REVERSE TRANSCRIPTION
Total cellular RNA was extracted from frozen tissues and cultured cells with Trizol reagent (Life Technologies, Cergy Pontoise, France) according to the manufacturer's instructions.
The reverse transcription reaction and PCR amplification were performed in a Crocodile III thermal reactor (Appligene, Illkirch, France). Aerosol resistant tips were used to prevent contamination. Total RNA (5 μg) was heated at 72°C for three minutes and then cooled on ice. It was combined with 0.5 μg of random hexamers (Promega, Lyon, France), transcription buffer (50 mM Tris HCl, pH 8.3, 75 mM KCl, 3 mM MgCl2, 10 mM DTT), dNTPs (1 mM each), RNasin (50 units; Promega), and RNaseH− M-MLV reverse transcriptase (200 units; Promega) in a final volume of 20 μl. Incubation was performed at 42°C for 60 minutes.
SEMIQUANTITATIVE EXPRESSION OF THECD155 GENE
Amplifications were performed with 3 μl of cDNA in a total volume of 50 μl containing 10 mM Tris HCl, pH 9.0, 50 mM KCl, 1.5 mM MgCl2, 0.1% Triton X-100, 0.2 mg/ml gelatin, 200 μM of each dNTP, 100 pmol of specific primers, and 2.5 units of Taq DNA polymerase (Promega).
Primers were designed from the published sequence of the humanCD155 gene.10 They were designed to amplify cDNA but not contaminating genomic DNA. The primers used to quantify expression of the CD155gene were located in a region corresponding to the extracellular domain of the protein, which is conserved in all splice variants (fig 1). The sequences of these primers were: CD1–5′ primer: 5′-TATCTGGCTCCGAGTGCTT GCC-3′; CD2–3′ primer: 5′-ATCATAGCCA GAGATGGATACC-3′.

Overview of the different CD155 transcripts and localisation of the oligonucleotides used for reverse transcription-polymerase chain reaction (RT-PCR) analysis. (A) Exon/intron organisation of the CD155 gene. Oligonucleotides CD1 and CD2, used to quantify expression of the CD155 gene, are located in a region corresponding to the extracellular domain of the protein which is conserved in all splice variants. Primers CD5 and CD15, used for qualitative analysis of CD155 expression, are located on each side of the different splice sites. Part of intron 6 is missing in variant β. The entire exon 6 is missing from variant γ. The predicted size of the different amplicons is indicated. (B) Agarose gel electrophoresis of CD155 transcripts amplified by RT-PCR from SW1116 colon cancer cells. Amplification was performed with oligonucleotides CD1 and CD2 (lane 1) or CD5 and CD15 (lane 2). No DNA was amplified when the reverse transcription was performed in the absence of reverse transcriptase.
For semiquantitative amplification, each cycle was carried out at 92°C for 30 seconds, 62°C for 30 seconds, and 72°C for 60 seconds. The reaction was stopped after 18 cycles, in the linear range of the amplification (not shown). Following electrophoresis, amplified DNA was transferred to Hybond N+ membranes (Amersham Pharmacia Biotech, Orsay, France). The nylon filter was hybridised using alkaline phosphatase labelled probes using the AlkPhos system (Amersham Pharmacia Biotech). For CD155, a 362 bp insert amplified by PCR from HCT8R cell cDNA with oligonucleotides CD1 and CD2 was used as a probe.
Normalisation was performed by RT-PCR amplification and hybridisation of β-actin cDNA. Fourteen cycles (92°C for 30 seconds, 58°C for 30 seconds, 72°C for 30 seconds) were performed with oligonucleotides B1 (5′-CCTTCCTGGGCATGGAGTCCT-3′) and B2 (5′-GGAGCAATGAT-CTTGATCT TC-3′). The amplified products (202 bp) were hybridised as described for CD155.
QUALITATIVE ANALYSIS OF CD155 EXPRESSION
For analysis of CD155 variants, 35 cycles of PCR were performed as described above. The primers used were located on exons 5 and 7, respectively, on regions corresponding to each side of the transmembrane domain, where the different splice sites are located (fig 1A). The sequences of these primers were: CD5–5′ primer: 5′-TCCTGTGGACAAACCAATCA ACAC-3′; CD15–3′ primer: 5′-GAGGCGCT GGCATGCTCTGT-3′.
Amplified fragments were identified by size after electrophoresis on a 3% agarose gel stained with ethidium bromide (fig 1B). For quantification, the bands of interest were stored as TIFF files using the BioCapt software (Vilber Lourmat, Marne la Vallée, France) and densities were determined using the EasinWin 32 software (Herolab, Fischer Scientific, Illkirch, France). The relative ratio of the different amplicons was determined for each reaction and expressed as a percentage of the overall expression. All samples were analysed at least three times.
Results
CD155 mRNA EXPRESSION IN PRIMARY CRC BY RT-PCR ANALYSIS
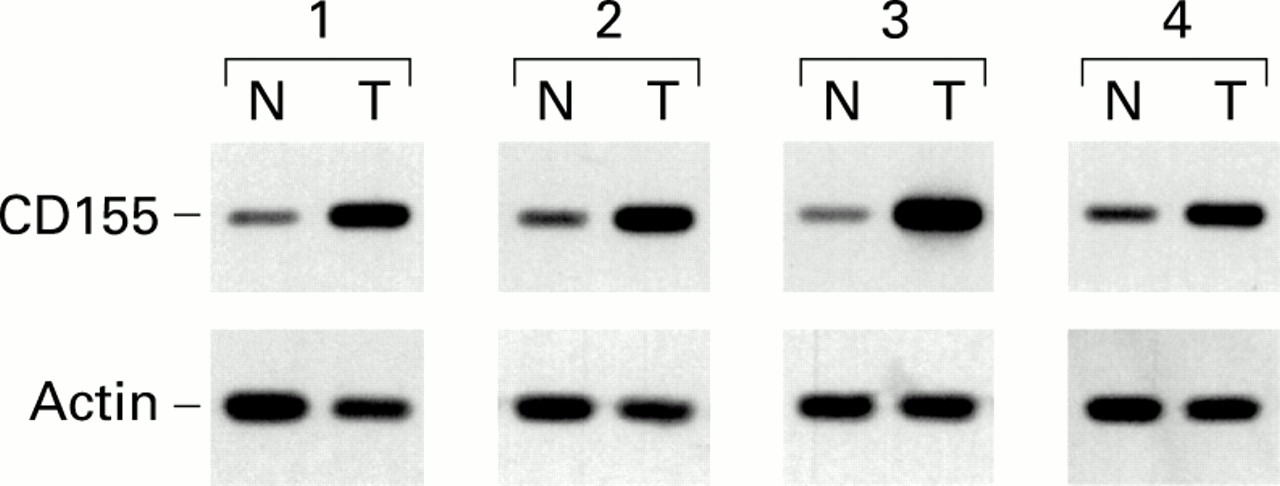
Firstly, we analysed expression of the humanCD155 gene in human colorectal tumours by RT-PCR and DNA hybridisation. RNA was extracted from six different tumours and the corresponding adjacent normal mucosa. Semiquantitative RT-PCR analysis was performed. Expression of theCD155 gene was higher in all tumour samples compared with adjacent normal tissues, as illustrated in fig 2. Quantification of the signals obtained indicated that there was a 2–10-fold increase in amounts of CD155 mRNA in colorectal tumours compared with normal mucosa.

Increased levels of CD155 in human colorectal carcinoma. Reverse transcription-polymerase chain reaction analysis of CD155 expression was performed on paired tumorous (lanes T) and non-tumorous (lanes N) colorectal tissues from four patients, as described in materials and methods.
CD155 EXPRESSION IN PRIMARY CRC BY IMMUNOHISTOCHEMISTRY
To determine which cells overexpress theCD155 gene, immunostaining experiments were performed on frozen 5 μm thick sections with monoclonal antibody D171. As illustrated in fig 3A, anti-CD155 reactivity was clearly detected in tumour samples. In each case the sections used for the negative control, which were stained with an irrelevant antibody, were negative (fig 3B). CD155 expression was localised to tumour cells and not observed in the stromal reaction under these experimental conditions. Similar staining of CD155 was identified in all adenomas as well as in all adenocarcinomas. Staining was independent of tumour grade, degree of differentiation, and localisation of the tumour. All normal colon tissues presented only weak staining of epithelial cells, and no staining was detected in the lamina propria (fig3C).

{kind=link}
{kind=link}
{kind=link}
CD155 is overexpressed in malignant epithelial cells in colorectal carcinoma. (A) Colon adenocarcinoma shows immunoreactivity with monoclonal antibody D171 (×200). (B) Adjacent section stained with an irrelevant monoclonal antibody (×200). (C) Normal colonic tissue showing barely detectable CD155 immunoreactivity (×200). (D) Normal section stained with an irrelevant monoclonal antibody (×200).
CD155 EXPRESSION IN HUMAN CANCER CELL LINES
In addition to the immunohistochemical staining experiments, D171 reactivity was observed by flow cytometry and indirect immunofluorescence with all human colon carcinoma cell lines tested so far (HT29, Colo205, SW620, HCT8R, SW1116; not shown).
CD155 EXPRESSION IN CRC BY IMMUNOBLOT ANALYSIS
To characterise further the protein detected by anti-CD155 monoclonal antibodies in human tissues, we performed western blot analyses. Proteins were extracted from four colon tumours and three colon cancer cell lines. Monoclonal antibody D171 is not suitable for western blotting. Therefore, following electrophoresis and transfer, nitrocellulose membranes were probed with monoclonal antibody 5H5 (kindly provided by Dr A Nomoto) which recognises CD155 under denaturing conditions (unpublished). This revealed an ∼80 kDa protein (data not shown) which is in agreement with the reported molecular weight of the transmembrane CD155 protein.14
EXPRESSION OF CD155 SPLICING VARIANTS
The CD155 gene has been shown to be expressed as several isoforms corresponding to splicing variants.10 To analyse the relative expression of these different transcripts in CRC, a qualitative RT-PCR analysis was performed with oligonucleotides CD5 and CD15. These primers are located on exons 5 and exon 7, respectively. Agarose gel electrophoresis revealed three amplified DNA fragments (fig 1B). These different fragments were purified, cloned, and sequenced. This analysis revealed that the major band (273 bp) corresponds to the isoform containing the transmembrane domain (isoform α). The lower 137 bp and 113 bp fragments correspond to transcripts lacking part of or the entire exon 6, respectively. They have been described as β and γ isoforms by Koike and colleagues.10 They lack the transmembrane domain and code for soluble forms of CD155.
cDNAs prepared from RNA extracted from tumours and normal tissues were analysed using this assay. The relative abundance of these three RNA transcripts was determined for six colon tumours and the corresponding normal mucosa. This revealed no qualitative difference between the samples studied (not shown).
All of these samples contain different cell types. Therefore, we applied this assay to preparations of normal intestinal epithelial cells isolated from surgical specimens obtained from patients undergoing colonic resection and compared the results with those obtained with human colon carcinoma cells lines. In normal epithelial cells, the transmembrane α isoform represents the main CD155 transcript (65–70%) (table 1). Variants β and γ correspond to 25–30% and 5–8% of the CD155 mRNAs, respectively. No significant difference was observed between the three preparations tested. Similar ratios between the different splicing variants were obtained with the colon cancer cell lines analysed (table 1). These data indicate that overexpression of the CD155 gene does not involve a particular isoform but most likely results from increased transcription of the gene.
Relative expression (%) of the different CD155 variants in normal epithelial cells and colon cancer cells
Discussion
The CD155 gene codes for integral membrane or soluble proteins that are members of the Ig superfamily.15 They share a V-C2-C2 domain structure.7 ,10 The members of this family have a diversity of functions but in most cases the common denominator is a recognition role at the cell surface.15 Most of these proteins are involved in cell-cell interaction during normal or pathological processes such as embryogenesis (for example, neural cellular adhesion molecule (N-CAM), and neurone-glia CAM (Ng-CAM)), immune response (Igs, T cell receptors, MHC antigens), inflammation and wound healing (intercellular CAM-1 (ICAM-1), CD54; vascular CAM-1 (VCAM-1), CD106), tumorigenesis and cancer metastasis (N-CAM; Ng-CAM; VCAM-1; ICAM-1; platelet endothelial CAM-1 (PECAM-1), CD31; carcinoembryonic antigen (CEA)). Some growth factor receptors also belong to the Ig family: the platelet derived growth factor receptor, colony stimulating factor 1 receptor, and fibroblast growth factor receptors. Finally, several viral infections are mediated through binding to a membrane receptor belonging to the Ig family.16
Human poliovirus infections are mostly targeted to the gastrointestinal tract through binding to the transmembrane protein coded by theCD155 gene.7 ,14 This is the only known function of the CD155 protein at present but it probably has important biological activities. Indeed, CD155 belongs to a small subfamily of the Ig superfamily that includes nectin 1, nectin 2, and nectin 3, all three of which have been shown to mediate cell-cell adhesion and to localise to sites of cadherin based cell junctions.17-19
As it is overexpressed in cancer cells, it is tempting to speculate that CD155 expression may play a role in neoplastic transformation and/or progression. CD155 overexpression was seen in all adenomas which are established premalignant lesions in colorectal carcinogenesis. Thus CD155 expression may play a role at early stages of tumorigenesis.
Several members of the Ig family are overexpressed in cancer cells. The CEA and NCAgenes are overexpressed in human CRC.20 ,21 The humanB-CAM gene is highly expressed in ovarian carcinomas in vivo.22 Muc18 has been identified as a marker of tumour progression in human malignant melanoma that may have a role in the development of metastasis.23 Finally, de novo expression of ICAM-1, the major human rhinovirus receptor,24 ,25 correlates with increased risk of metastasis in melanoma.26
The qualitative RT-PCR that we have developed revealed that the relative abundance of the three RNA transcripts was not significantly different between cancer cells and normal epithelial cells. This indicated that there is no difference between RNA splicing mechanisms in normal and colon cancer cells but rather that there is increased transcription of the CD155 gene in colon cancer cells.
The promoter of the human CD155 gene has been isolated.27 Solecki and colleagues28recently reported mapping of three ciselements within the core promoter, and demonstrated that the AP-2 transcription factor is involved in transcription of theCD155 gene in HeLa cervical carcinoma cells. Further investigation into the regulation of theCD155 gene in colorectal cancer cells is necessary. We are also investigating the potential use of soluble CD155 molecules as tumour markers. In addition, demonstration that theCD155 gene is overexpressed at early stages of colorectal carcinogenesis should initiate studies on the function(s) of the different CD155 isoforms.
Finally, during completion of our manuscript, Gromeier and colleagues29 demonstrated that highly attenuated poliovirus recombinants can infect and propagate in cell lines derived from malignant gliomas and, most interestingly, halted tumour progression and eliminated tumours in athymic mice. They proposed that susceptibility of these malignant cells to poliovirus may be mediated by expression of the CD155 gene in glial neoplasms. Our demonstration that the CD155gene is overexpressed in colorectal cancer cells suggests that oncolytic poliovirus recombinants may also be suitable for the treatment of colorectal cancer.
Acknowledgments
This work was supported by a grant from the Ligue Départementale (44) Contre le Cancer. B Baury is the recipient of a research fellowship from the Ligue Départementale (44) Contre le Cancer. We thank Dr Akio Nomoto (Department of Microbiology, Tokyo University, Japan) for providing the monoclonal antibodies directed against human CD155. A Jarry and B Baury contributed equally to this work.
Abbreviations used in this paper
- CRC
- colorectal carcinoma
- RT-PCR
- reverse transcription-polymerase chain reaction
- CAM
- cellular adhesion molecule
- CEA
- carcinoembryonic antigen
- Ig
- immunoglobulin