Article Text

Abstract
Background and aims: Inflammatory bowel diseases (IBDs) are associated with uncontrolled innate and adaptive immunity against normal constituents, including commensal bacteria and microbial products. Mesenchymal stem cells (MSCs) suppress effector T cell responses and have beneficial effects in various immune disorders. This work investigates the therapeutic effects of human adipose-derived MSCs (hASCs) in various models of IBD and sepsis.
Methods: Acute and chronic colitis was induced in mice with dextran sulfate sodium. Sepsis was induced by caecal ligation and puncture or by endotoxin injection. Colitic and septic mice were treated intraperitoneally with hASCs or murine ASCs, and diverse disease clinical signs and mortality were determined. The levels of various inflammatory cytokines and chemokines, T helper 1(Th1)-type response and generation of regulatory T cells (Treg) were determined in affected organs.
Results: Systemic infusion of ASCs significantly ameliorated the clinical and histopathological severity of colitis, abrogating weight loss, diarrhoea and inflammation, and increasing survival. The therapeutic effect was associated with downregulation of the Th1-driven inflammatory responses. ASCs decreased a wide panel of inflammatory cytokines and chemokines and increased interleuklin 10 (IL10), acting on macrophages. hASCs also impaired Th1 cell activation in both colonic mucosa and draining lymph nodes. The induction of IL10-secreting Treg was partially involved in the therapeutic effect of hASCs. Moreover, ASCs protected from severe sepsis by reducing the infiltration of inflammatory cells in various target organs and by downregulating the production of various inflammatory mediators.
Conclusions: hASCs emerge as key regulators of immune/inflammatory responses in vivo and as attractive candidates for cell-based treatments for IBD and sepsis.
Statistics from Altmetric.com
Inflammatory bowel disease (IBD), including Crohn’s disease (CD) and ulcerative colitis, is a family of chronic, idiopathic, relapsing and tissue-destructive diseases characterised by dysfunction of mucosal T cells and altered cellular inflammation that ultimately leads to damage of the distal small intestine and the colonic mucosa.1 In both CD and experimental colitis, activated T helper 1 (Th1) cells promote an exaggerated macrophage and neutrophil infiltration and activation, giving rise to a transmural inflamed intestinal mucosa, characterised by prolonged and uncontrolled production of inflammatory cytokines and chemokines.1 2 Cytokines and free radicals produced by infiltrating cells and resident macrophages play a critical role in colonic tissue destruction. The fact that most encounters with mucosal antigens do not generate effector T cell responses, but instead induce suppressive regulatory T cell (Treg) responses, anergy or deletion of antigen-specific T cells, links CD to a failure of the mucosal immune system to attenuate the immune response to endogenous antigens.1 3 Recent evidence supports the notion that an imbalance between effector T cells and Treg, probably by a defect in the Treg repertory, results in expansion of self-reactive T cells and inflammation.3 The failure in the current treatments for IBD illustrates the need for novel therapeutic approaches that specifically modulate both components of the disease—that is, the inflammatory and T cell-driven responses—and try to restore immune tolerance by re-establishment of the Treg repertory.
Beside their capacity to differentiate into various cell lineages and their clinical interest in tissue repair,4 mesenchymal stem cells (MSCs) have emerged as potent regulators of the immune response. Thus, bone marrow-derived MSCs (BM-MSCs) inhibit in vitro T cell activation, B cell function and dendritic cell (DC) maturation, and efficiently protect against allograft rejection and experimental autoimmunity.5–14 However, a critical issue for clinical translation of BM-MSCs in immune disorders is that large quantities of infused cells are required for treatment, which in most cases are not available. Human MSCs obtained from subcutaneous adipose tissue (hASCs) have lately emerged as an attractive alternative MSC source for cell treatment.15 Large amounts of hASCs can be easily obtained from lipoaspirates of healthy donors and rapidly expanded in vitro to generate a clinically effective dosage. Moreover, hASCs share some of the immunomodulatory properties that characterise the BM-MSCs.16–18 Indeed, murine ASCs (mASCs) were effective in preventing graft-versus-host disease.16 Here, we characterise the immunosuppressant activity of hASCs and mASCs in xenogeneic, allogeneic and syngeneic experimental models of IBD and sepsis and their capacity to inhibit the inflammatory response in vivo.
METHODS
Cell preparation
hASCs and mASCs were obtained, characterised and cultured as described in Supplementary methods.15 16
Induction of acute and chronic colitis
Acute colitis was induced in C57Bl/6 mice (7-week-old, Charles River) by administering 5% dextran sulfate sodium (DSS; molecular weight 40 000 Da; Sigma, St Louis, Missouri, USA) from day 0 to day 7 in the drinking water ad libitum. At day 2, hASCs (105–5×106), syngeneic mASCs (106, from C57Bl/6 mice) or allogeneic mASCs (106, from Balb/c mice) were injected intraperitoneally in the DSS-treated animals. Chronic colitis was induced in C57Bl/6 mice by administering 3% DSS in the drinking water in a cyclic manner,19 which consisted of two cycles of 7 days with DSS, followed by a 10-day period without DSS supplementation. On day 7 of each cycle, mice were injected intraperitoneally with hASCs (106 cells/mouse). Colitis severity was assessed daily by scoring (0–4) the clinical disease activity by evaluating stool consistency, presence of faecal blood and weight loss.19 Mice with acute colitis were sacrificed on day 5 and mice with chronic colitis were sacrificed on days 10 or 26. The entire colon was removed from the caecum to the anus, and colon length and weight were measured as indirect inflammation markers. The macroscopic colonic damage score was assessed based on the grade of tissue adhesion, presence of ulceration and wall thickness.19 Colon segments were processed for histopathological analysis or frozen in liquid nitrogen for protein extraction, and cytokine and myeloperoxidase (MPO) activity measurements as described in Supplementary methods. To trace the injected cells in vivo, hASCs were labelled with 5,6-carboxyfluorescein succimidyl ester (CFSE) before injection and analysed as described in Supplementary methods.
Induction of endotoxaemia and sepsis
To induce endotoxaemia, Balb/c mice were injected intraperitoneally with lipopolysaccharide (LPS; 400 μg/mouse). To induce sepsis, caecum of anesthetised Balb/c mice was ligated 5.0 mm from the caecal tip, punctured once with a 22-gauge needle and the stool then extruded (1 mm). Mice were treated intraperitoneally with medium or hASCs (105–106 cells) 30 min after LPS injection or 4 h after caecum perforation, and monitored for survival. Serum, peritoneal exudates, liver, lungs and small intestines were collected at different times for protein extraction, and cytokine and MPO activity determinations.
All experiments were performed according to the Institutional Guidelines for the Care and Use of Laboratory Animals in Research and approved by Consejo Superior de Investigaciones Cientificas.
Mesenteric lymph node (MLN) cell cultures
Single-cell suspensions (106 cells/ml) obtained from MLNs at the disease peak were incubated in complete medium (RPMI-1640 supplemented with 100 U/ml penicillin/streptomycin, 2 mM l-glutamine, 50 μM 2-mercaptoethanol, 10% fetal calf serum (FCS)) without (unstimulated) or with phorbol 12-myristate 13-acetate (PMA; 10 ng/ml) and concanavalin A (Con A; 2.5 μg/ml). Cell proliferation (expressed as A450) was evaluated for 96 h by using a cell proliferation 5-bromodeoxyuridine (BrdU) assay (Roche, Kulmbach, Germany). Cytokine/chemokine secretion and intracellular cytokine contents were determined after 48 h culture as described in the Supplementary methods.
Statistical analysis
All results are expressed as mean (SD). The non-parametric Mann–Whitney U test was applied on all clinical results and cell culture experiments. Changes in the body weight were compared by using the Wilcoxon matched-pair signed-rank test. Survival was analysed by the Kaplan–Meier log-rank test. p<0.05 was considered significant.
RESULTS
hASCs show potent immunomodulatory activities
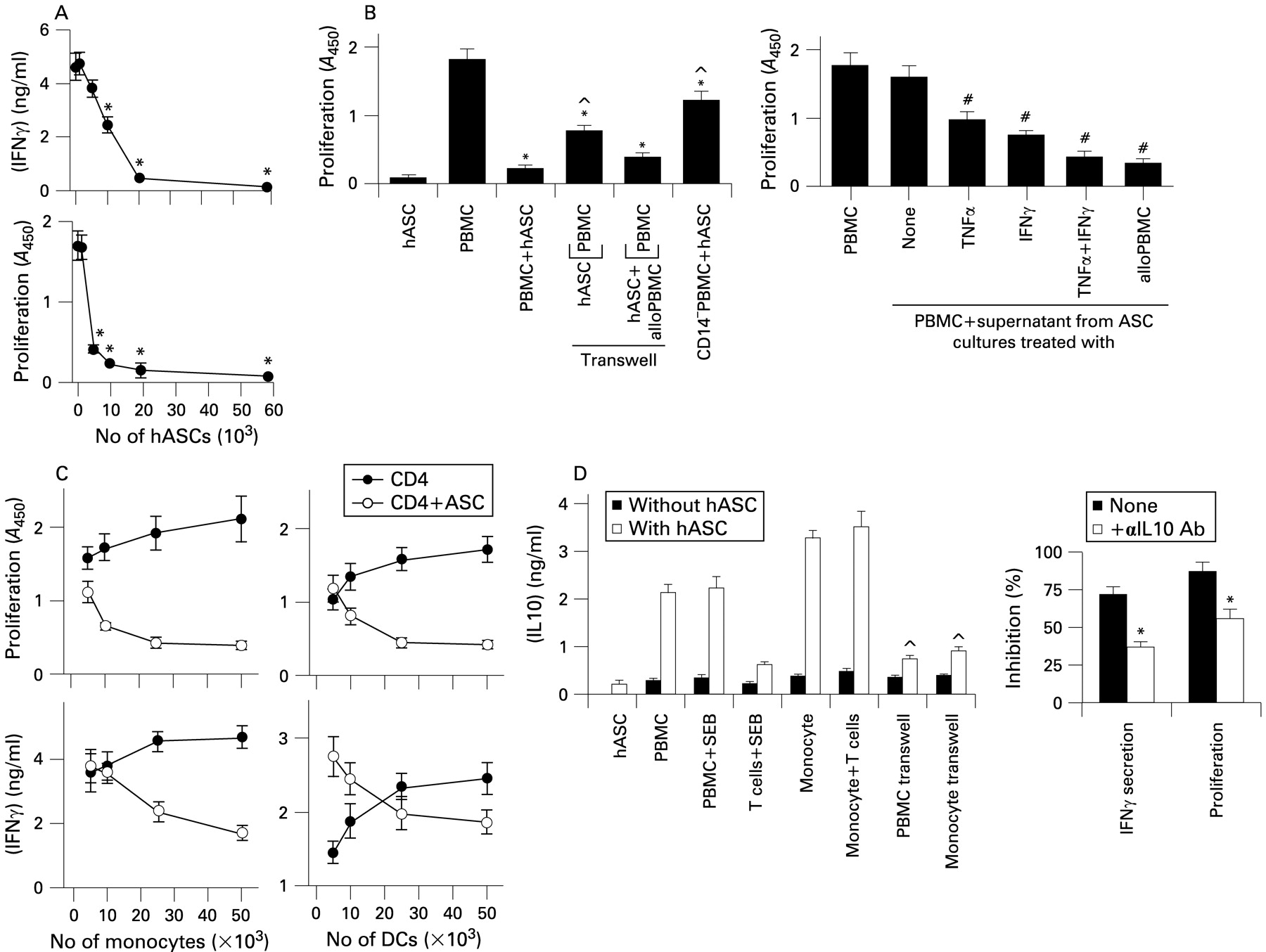
Recent studies characterised the phenotype of hASCs and their immunoregulatory effects on T cells.16–18 We first confirmed the ability of hASCs to inhibit T cell activation with the superantigen staphylococcal enterotoxin E (SEB), as measured by cytokine secretion and T cell proliferation (fig 1A). The inhibitory effect was partially reversed when peripheral blood mononuclear cells (PBMCs) and hASCs were separated by a semi-permeable transwell membrane (fig 1B), suggesting a partial cell–cell contact dependence. The partial involvement of soluble factors produced by hASCs was supported by the fact that conditioned media from activated hASCs suppressed SEB-induced proliferation of PBMCs (fig 1B). Moreover, the co-culture of allogeneic PBMCs and hASCs in the same chamber of the transwell system fully restored their inhibitory activity on SEB-activated PBMCs situated in the other chamber (fig 1B), suggesting that PBMC–hASC contact induces the secretion of an immunosuppressive factor(s) for T cells. Interestingly, depletion of CD14 cells from PBMCs significantly abrogated the suppressive effect of hASCs (fig 1B), suggesting a partial indirect involvement of monocytes in such an effect. Moreover, addition of increasing numbers of monocytes or DCs into hASC–CD4 T cell co-cultures progressively inhibited proliferation and interferon γ (IFNγ) production (fig 1C). These data support a pivotal role for antigen-presenting cells (APCs) on the suppressive activity of hASC, and also that the inhibitory signal from the hASCs toward APCs depends on cell–cell contact. Because interleukin 10 (IL10) is a potent immunosuppressive cytokine produced by APCs, we investigated the involvement of IL10 in this system. IL10 production increased, in a cell–cell contact-dependent manner, in co-cultures of hASCs with PBMCs or monocytes, but not with T cells (fig 1D). IL10 blockade partially reversed the inhibitory activity of hASCs on T cell activation (fig 1D).

Immunomodulatory properties of human adipose-derived mesenchymal stem cells (hASCs). (A) hASCs inhibit peripheral blood mononuclear cell (PBMC) activation. Human PBMCs were stimulated with staphylococcal enterotoxin E (SEB) in the absence or presence of increasing numbers of hASCs. The proliferative response was determined after 96 h culture. Interferon γ (IFNγ) contents in the supernatants were determined after 72 h culture. Results are the mean (SD) of four experiments performed in duplicate. *p<0.001 vs SEB-stimulated PBMCs. (B) The inhibitory activity of hASCs depends on both cell–cell contact and soluble factors. Left panel: PBMCs were stimulated with SEB in the absence or presence of hASCs. When indicated, PBMCs were stimulated with SEB in the upper chamber of a transwell system separated from hASCs or hASCs plus allogeneic PBMCs (alloPBMC; situated in the lower chamber) by a semi-permeable membrane, and proliferation determined in the upper chamber. In some experiments, PBMCs were depleted of CD14 cells before the co-culture with hASCs. Right panel: hASCs were cultured with medium (none), tumour necrosis factor α (TNFα), IFNγ or allogeneic SEB-activated PBMCs (alloPBMC). After 24 h, culture supernatants were collected and added to PBMC cultures stimulated with SEB, and proliferation was determined after 96 h. Results are the mean (SD) (n = 4). *p<0.001 vs SEB-stimulated PBMCs. ^p<0.001 vs PBMC–ASC co-cultures. #p<0.001 vs SEB-stimulated PBMCs. (C) Immunosuppression by hASCs is partially mediated through monocytes and dendritic cells (DCs). CD4+ T cells were stimulated with SEB in the absence or presence of hASCs and increasing numbers of either monocytes (left panels) or DCs (right panels). T cell activation was followed by either IFNγ secretion or proliferation. Results are the mean (SD) of four experiments performed in duplicate. (D) hASCs induce the production of interleukin 10 (IL10) by monocytes through a cell–cell contact mechanism. T cells (5×104), monocytes (5×104), PBMCs (105) and hASCs (2×104) were cultured alone or in combination, in the absence or presence of SEB. When indicated, hASCs were separated from monocytes or PBMCs in a transwell system. After 48 h, culture supernatants were assayed for IL10 secretion by ELISA. The right panel represents the percentage of inhibition of IFNγ secretion and of the proliferative response of hASCs on SEB-activated PBMCs in the absence (none) or presence of IL10-neutralising antibodies (αIL 10 Ab). ^p<0.001 vs hASC–monocyte and hASC–PBMC co-cultures without transwell; *p<0.01 vs samples treated without anti-IL10 antibodies (n = 4).
hASCs protect against DSS-induced severe colitis
Once the immunosuppressive activity of hASCs in vitro was confirmed, we next investigated the potential therapeutic action of hASCs in two experimental models of chronic and acute colitis induced by oral DSS administration. In these models, intestinal inflammation results from the impairment of the intestinal epithelial cell barrier function by DSS, subsequent exposition of the submucosa to various luminal antigens (bacteria and food) and activation of the inflammatory cells involved in the innate immunity.
By 7 days, oral administration of 5% DSS resulted in a significant increase in the disease activity index, characterised by acute colitis, bloody diarrhoea and sustained weight loss resulting in 100% mortality (fig 2A–C). Macroscopic examination of colons showed striking hyperaemia and inflammation, revealed by a decreased colon length/weight ratio (fig 2D). hASC treatment dose-dependently increased survival rate, ameliorated body weight loss, and improved the wasting disease and colon inflammation (fig 2A–E). Histological examination showed that hASC treatment reduced the DSS-induced colonic transmural inflammation, mucin-producing goblet cell depletion, epithelial ulceration, focal loss of crypts and infiltration of inflammatory cells in the lamina propria (fig 2F). Moreover, hASC-treated animals showed decreased colonic MPO activity, reflecting less neutrophil infiltration (fig 2G). Initiation of the hASC treatment 1 day before DSS administration did not significantly improve the beneficial effects shown by hASC treatment on established colitis, and hASC injection 1 week before colitis induction showed no protective effects (Supplementary fig 1).

Treatment with human adipose-derived mesenchymal stem cells (hASCs) protects against dextran sulfate sodium (DSS)-induced acute colitis. Mice received 5% DSS with their drinking water from day 0 to day 7. hASCs (106/mouse) were infused intraperitoneally on day 2. Disease activity scores (A), weight loss (B) and mortality (C) were determined daily. The dose–response effect of hASCs (105, 5×105, 106, 5×106/mouse) on disease activity was determined at day 7 (D). Colon length and weight (E) and histopathological signs (F) were determined at day 6. Neutrophil infiltration was determined by measuring myeloperoxidase (MPO) activity in colonic protein extracts obtained on day 6 (G). Mice receiving tap water instead of DSS were used as controls (naïve). n = 6–8 mice/group. *p<0.01 vs DSS-treated mice.
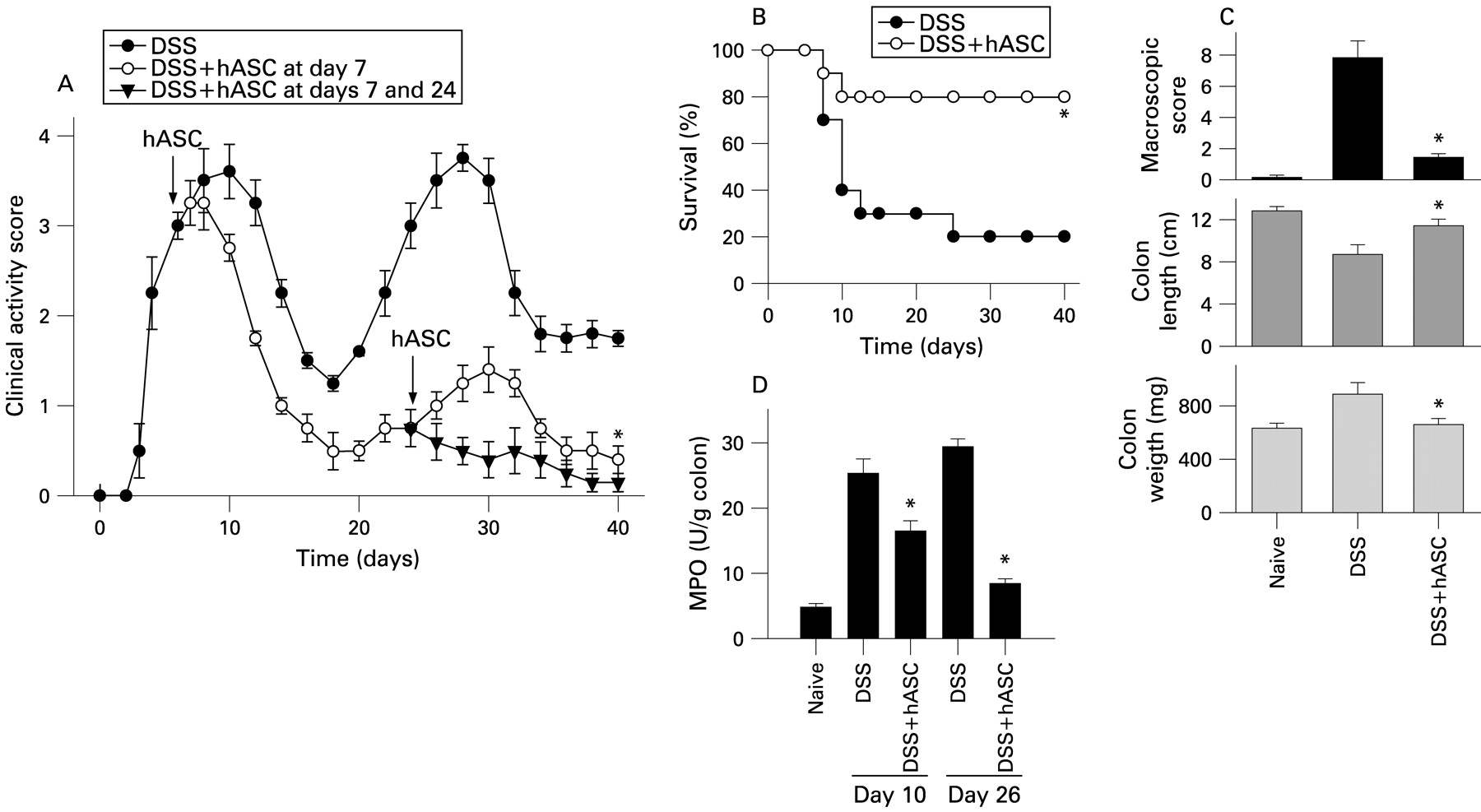
Administration of 3% DSS in two cycles resulted in two peaks of colitis characterised by continuous weight loss, diarrhoea and bloody stools, causing a mortality of 80% (fig 3A,B). Colitis was correlated with dramatic signs of colon damage and inflammation, characterised by colon shortening, increased organ weight and augmented colonic MPO activity (fig 3C,D). hASC injection during the first acute peak of colitis (day 7) significantly reduced the clinical activity by day 9, as shown by improvement of stool consistency, less rectal bleeding, decrease in colon shortening, reduced MPO activity, amelioration of colon damage and improvement in survival (fig 3A–D). Importantly, a single hASC infusion at the first acute colitis peak conferred significant resistance to disease activity during a second cycle of DSS administration (fig 3A). A second hASC injection during the second cycle of DSS infusion completely abrogated the clinical signs (fig 3A).

Treatment with human adipose-derived mesenchymal stem cells (hASCs) protects against dextran sulfate sodium (DSS)-induced chronic severe colitis. Mice received 3% DSS with their drinking water in a cyclic manner. Each cycle consisted of 7 days of DSS followed by a 10-day period without DSS supplementation. hASCs (106/mouse) were infused intraperitoneally on day 7 or on days 7 and 24 (arrows). The clinical activity score (A) and mortality (B) were determined daily. Macroscopic colonic damage score, colon length and colon weight were determined at day 26 (C). Neutrophil infiltration was determined by measuring myeloperoxidase activity in colonic protein extracts obtained on days 10 and 26 (D). Mice receiving tap water instead of DSS were used as controls (naïve). n = 6–8 mice/group. *p<0.001 vs DSS-treated mice.
The therapeutic effect of hASCs was not exclusive to the DSS-induced colitis model. hASC treatment was proven to be effective in ameliorating colitis induced with 2,4,6-trinitrobenzene sulfonic acid (TNBS; Supplementary Fig. 2).
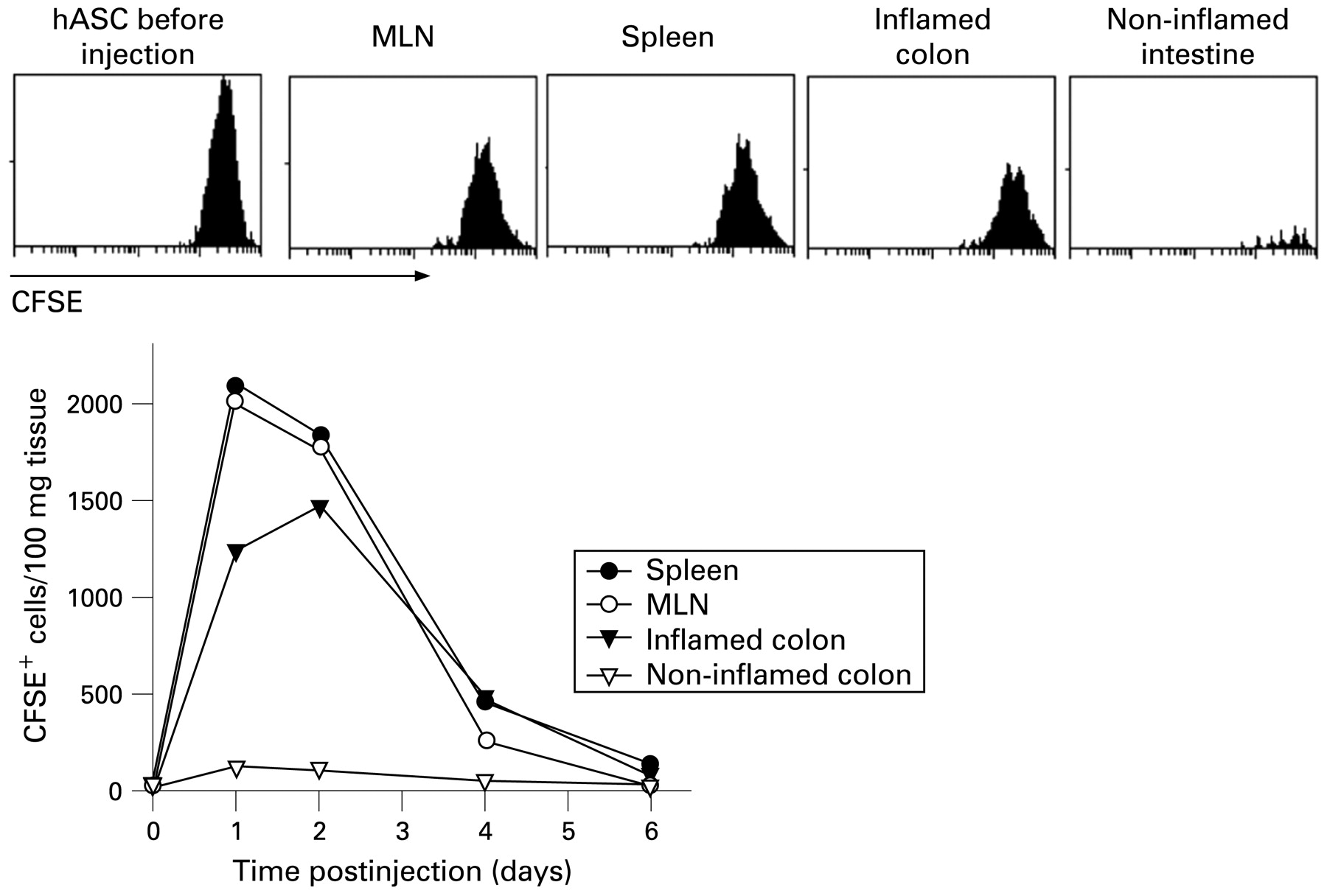
To understand better the half-life and trafficking of the infused hASCs, we injected CFSE-labelled hASCs into DSS-colitic mice. We detected the inoculated cells in the MLNs and spleen of the recipients 1–3 days postinjection (fig 4). Interestingly, hASCs were recruited by the inflamed colon, but not by non-inflamed intestine (fig 4). The specific homing of hASCs to secondary lymphoid organs and inflamed sites correlated with the expression by hASCs of a panel of chemokine receptors (Supplementary table).
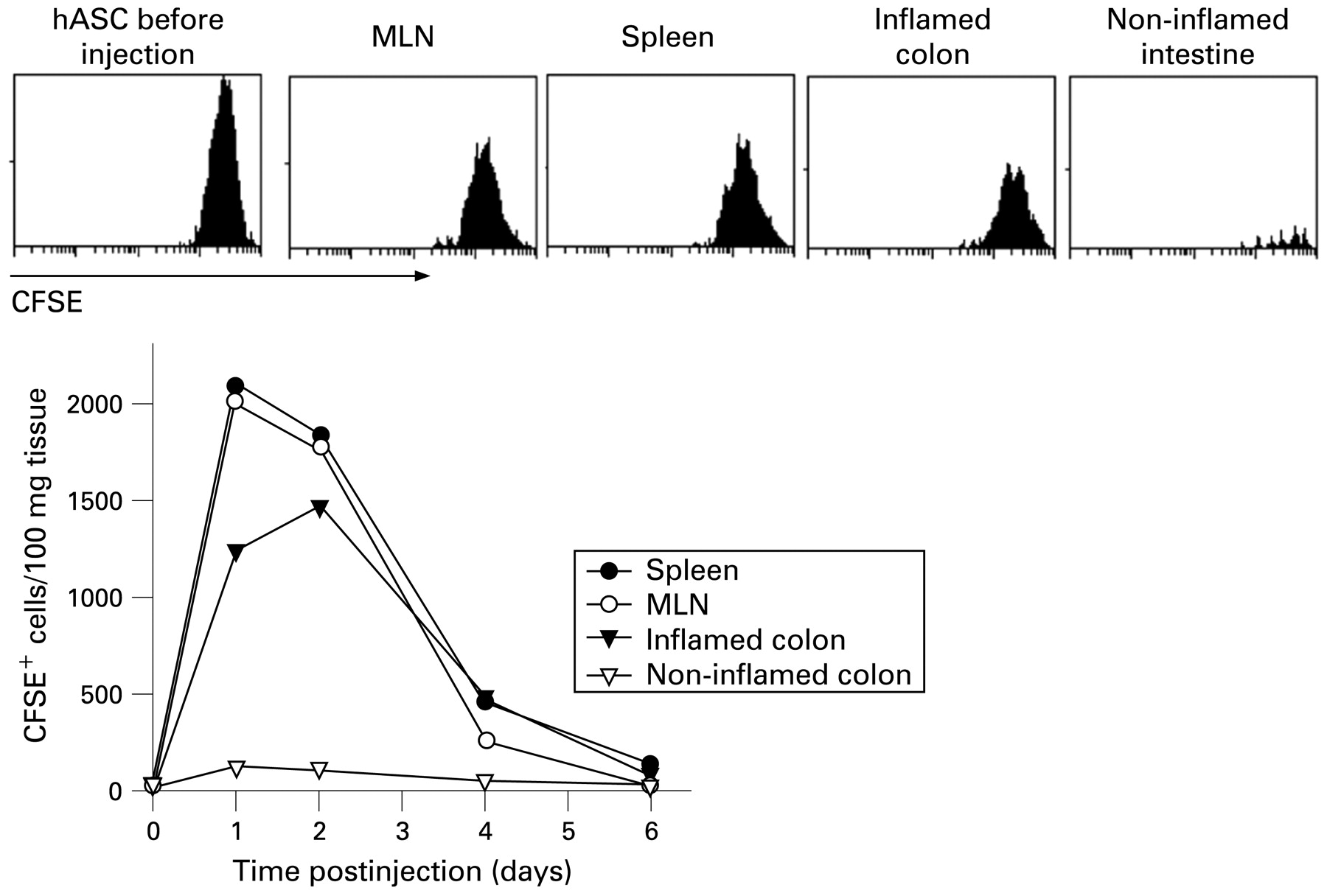
Human adipose-derived mesenchymal stem cells (hASCs) are preferentially recruited by lymphoid organs and inflamed colon. 5,6-Carboxyfluorescein succimidyl ester (CFSE)-labelled hASCs (106) were injected intraperitoneally on day 2 to dextran sulfate sodium (DSS)-colitic mice and their presence in various organs determined by flow cytometry at the indicated times after injection. Histograms correspond to samples obtained 36 h after hASC injection. The profile of CFSE-labelled hASCs before injection is included. Bottom panels represent the number of CFSE+HLA-ABC+CD11b− cells per 100 mg tissue. n = 3 mice/group. MLN, mesenteric lymph node.
hASC treatment reduces inflammatory responses in colitic mice
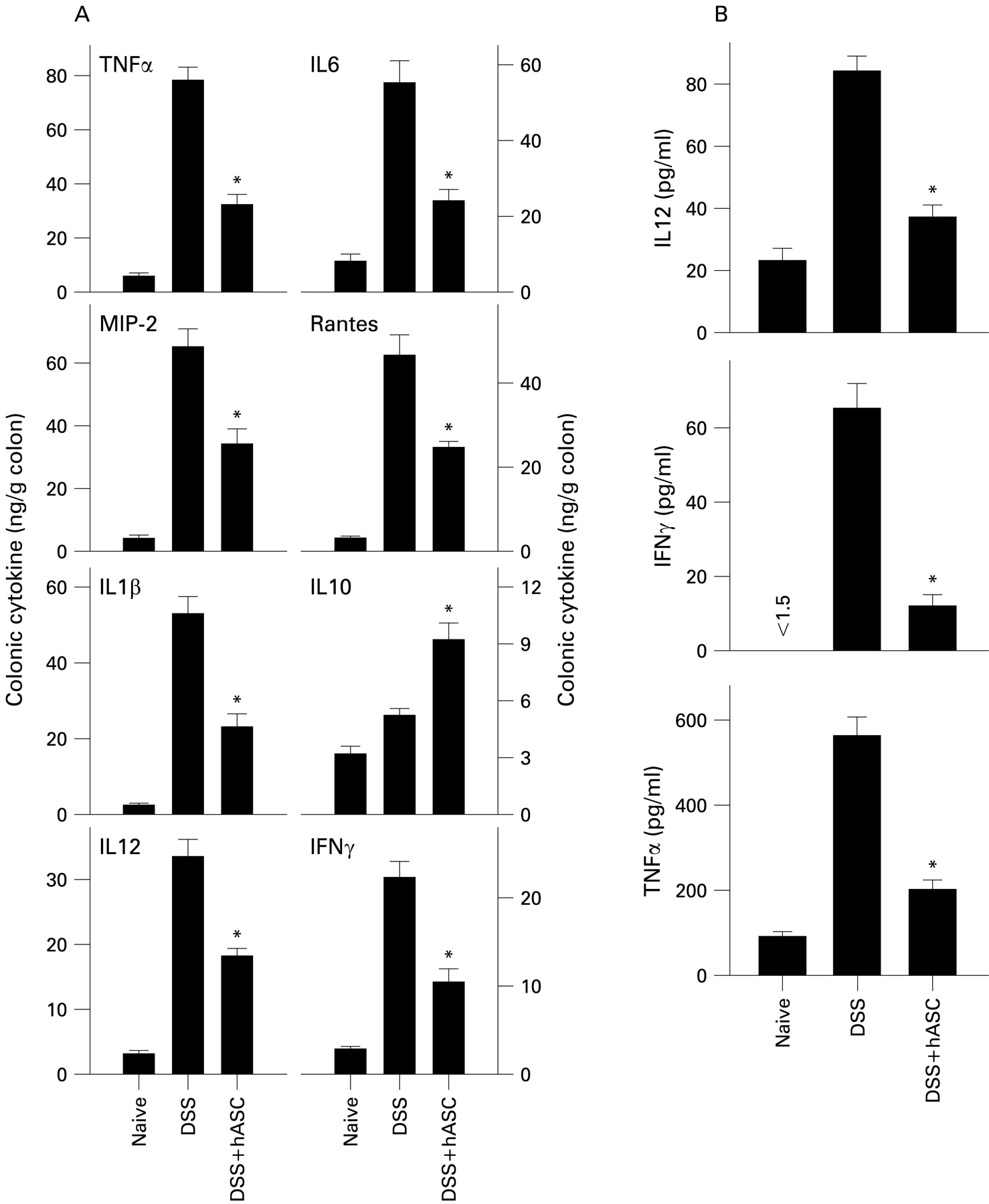
We next investigated the in vivo effect of hASCs on production of inflammatory mediators that are mechanistically linked to colitis. Colons of hASC-treated mice contained reduced levels of inflammatory cytokines (TNFα, IFNγ, IL6, IL1β and IL12) and chemokines (RANTES (regulated on activation normal T cell expressed and secreted) and macrophage inflammatory protein-2 (MIP-2)), and increased levels of the anti-inflammatory/regulatory cytokine IL10, in comparison with untreated DSS-colitic mice (fig 5A, Supplementary fig 3). This effect was not only a consequence of a diminished inflammatory infiltration in the mucosa, because mononuclear cells isolated from the lamina propria of hASC-treated mice produced less TNFα, IL12 and IFNγ on ex vivo culture (fig 5B), suggesting that hASCs deactivated the colonic inflammatory response.
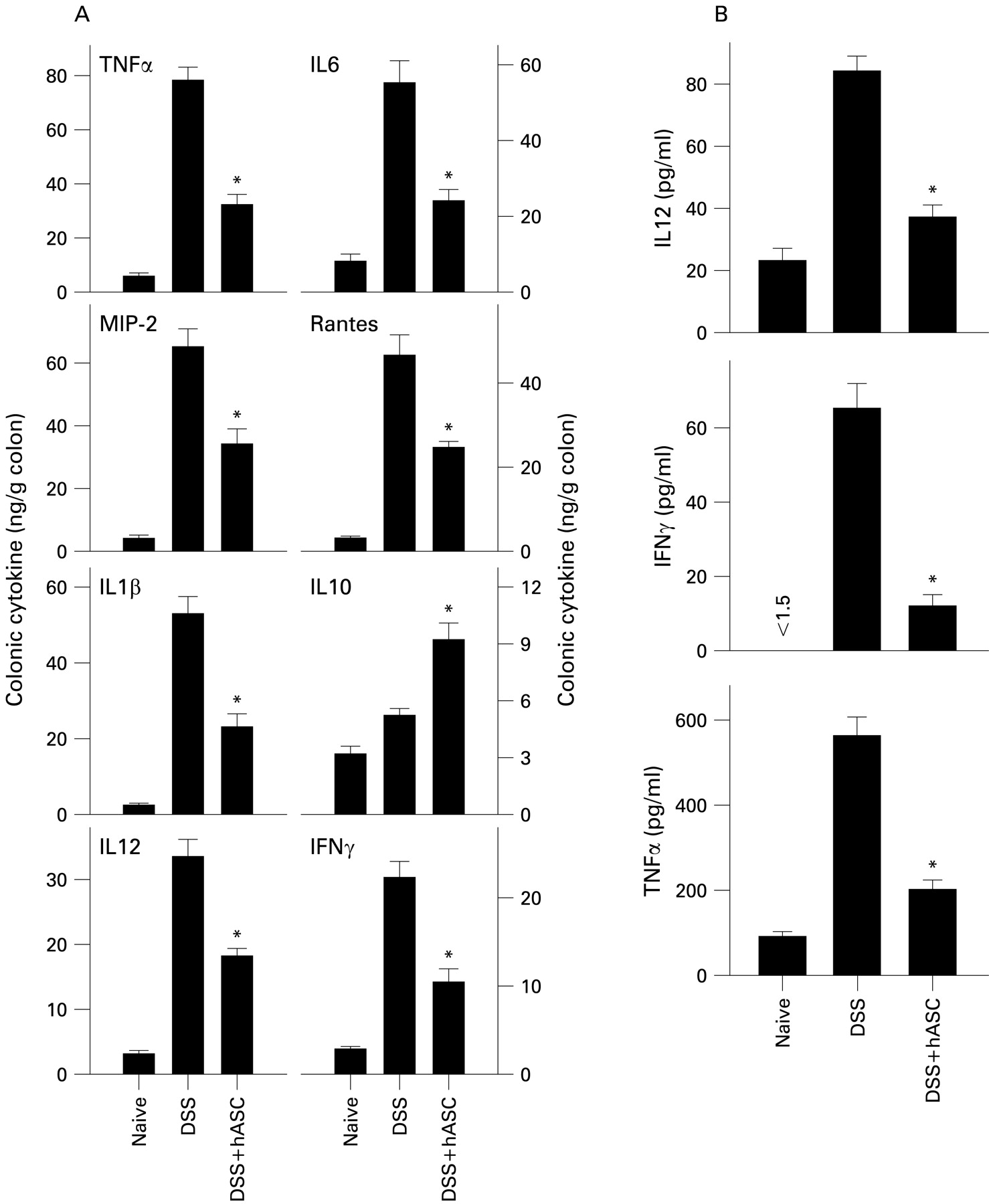
Treatment with human adipose-derived mesenchymal stem cells (hASCs) reduces colonic inflammatory responses in dextran sulfate sodium (DSS)-induced chronic colitis. Mice with DSS-induced chronic colitis were treated with hASCs at day 7. (A) Cytokine/chemokine contents in colonic protein extracts obtained on day 26 were determined. (B) Lamina propria cells were isolated at day 26, cultured without stimulation for 24 h and the cytokine contents determined in culture supernatants. n = 4 mice/group. *p<0.001 vs DSS-treated mice. IFNγ, interferon γ; IL, interleukin; MIP-2, macrophage inflammatory protein-2; RANTES, regulated on activation normal T cell expressed and secreted; TNFα, tumour necrosis factor α.
hASC treatment downregulates Th1-type responses and induces Treg in colitis
CD and DSS-induced colitis are generally considered to be Th1-mediated diseases, and the bias towards Th1 cytokines (mainly IFNγ and TNFα) is crucial in the establishment of chronic inflammation in the colon.1 2 We determined the ability of MLN cells isolated from hASC-treated colitic mice to proliferate and produce cytokines on ex vivo restimulation. DSS-induced inflammation in the gastrointestinal tract correlated with increased cellularity (mainly T cells and macrophages) of the MLNs, the major lymph nodes draining from the gut (fig 6A). Moreover, MLN cells obtained from DSS-treated mice showed marked proliferation and produced high IFNγ levels and low amounts of IL4 and IL10 (fig 6B). However, MLNs from hASC-treated colitic mice showed normalised MLN cellularity (fig 6A), proliferated much less and produced low levels of Th1 cytokines, even after ex vivo stimulation with PMA/Con A (fig 6B). The Th2-type cytokine IL4 was not affected (not shown), whereas the regulatory cytokine IL10 was significantly increased (fig 6B). Determination of intracellular cytokine expression in MLN CD4 cells showed that hASCs decreased the number of IFNγ-producing Th1 cells, and increased the number of IL10-producing CD4 T cells (fig 6C).

Treatment with human adipose-derived mesenchymal stem cells (hASCs) downregulates the T helper 1 (Th1) cytokine response and induces a regulatory T response in dextran sulfate sodium (DSS)-induced colitis. Mice with DSS-induced chronic colitis were treated with hASCs at day 7. (A) The number of mononuclear cells in mesenteric lymph node (MLN) cells isolated at day 10 was determined. (B) MLN cells were isolated at day 10 and cultured with medium alone (unstimulated) or with phorbol 12-myristate 13-acetate +concanavalin A (stimulated). The proliferative response was determined after 4 days culture. Cytokine contents in the supernatants were determined after 48 h of culture. (C) Intracellular expression of cytokines in CD4 T cells from MLNs isolated at day 10 was determined by flow cytometry. (D) The percentages and numbers of CD4+CD25+Foxp3+ cells in the CD4 population of MLNs isolated at days 10 and 26 were determined by flow cytometry. n = 4 mice/group. *p<0.001 vs DSS-treated mice. IFNγ, interferon γ; IL10, interleukin 10.
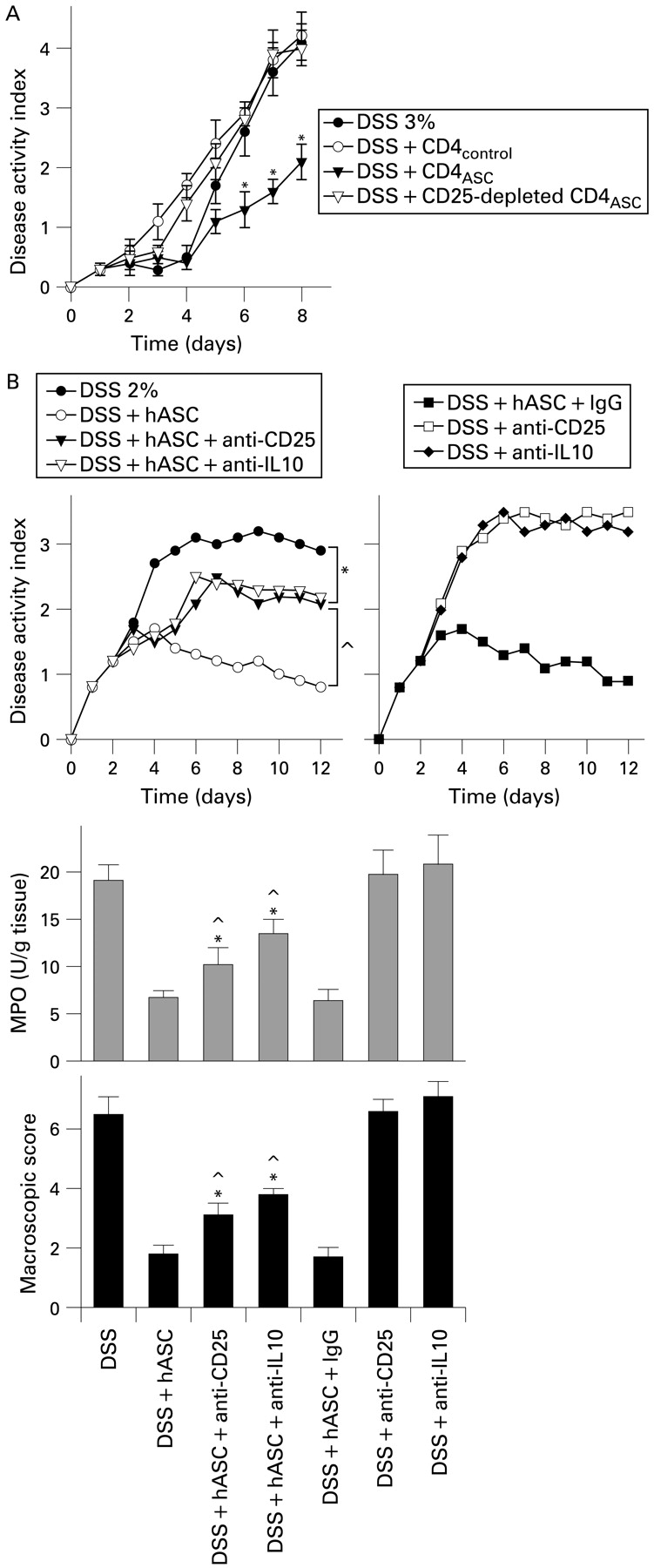
Several studies indicated that Treg confer significant protection against IBD by decreasing the activation and colon homing of autoreactive Th1 cells.20–22 The observed downregulation of the inflammatory/Th1 response and the elevated IL10 levels prompted us to investigate the capacity of hASCs to increase the Treg repertory in vivo. hASC-treated colitic mice had significantly higher numbers of CD4+CD25+Foxp3+ Treg in MLNs than untreated colitic mice (fig 6D), that persisted for a long period of time. To investigate the ability of these cells to inhibit the progression of colitis, DSS-colitic mice were treated with CD4 cells isolated from untreated (CD4control) or hASCs-treated (CD4ASC) colitic mice. In contrast to the CD4control, CD4ASC ameliorated colitis, and depletion of CD25+ cells from CD4ASC significantly abolished this therapeutic effect (fig 7A). In vivo depletion of IL10 or CD25+ T cells partially reversed the beneficial action of hASCs on colitis (fig 7B), demonstrating the involvement of IL10 and Treg in their therapeutic effect.

Partial involvement of T regulatory cells (Treg) and interleukin 10 (IL10) on the therapeutic effect of human adipose-derived mesenchymal stem cells (hASCs). (A) Effect of adoptive transfer of CD4+ and CD4+CD25− T cells isolated from mesenteric lymph nodes of untreated (CD4control) or hASC-treated dextran sulfate sodium (DSS) mice (CD4ASC) on the progression of DSS-induced acute colitis. n = 5 mice/group. *p<0.01 vs DSS-treated mice. (B) Mice with acute colitis were treated with hASCs and then depleted in vivo of IL10 or CD25+ T cells with neutralising antibodies. Injection of an immunoglobulin G (IgG) antibody was used as a control. Disease activity index was determined daily, and macroscopic score and myeloperoxidase (MPO) activity were determined at day 12. n = 4–5 mice/group. *p<0.01 vs untreated DSS-colitic mice. ^p<0.01 vs hASC-treated colitic mice. IgG, immunoglobulin G; MPO, myeloperoxidase.
Syngeneic and allogeneic mASCs protect against colitis
To investigate whether the beneficial effect observed for hASCs was a consequence of a treatment in a xenogeneic system, colitis was treated with syngeneic or allogeneic mASCs. Both allogeneic and syngeneic mASCs efficiently protected against DSS-induced acute colitis and mortality (fig 8A), and ameliorated the mucosal inflammatory response (fig 8B). Moreover, mASCs reduced in vivo the number of IL2/IFNγ-secreting T cells, while increasing the number of IL10-producing and CD25+FoxP3+CD4+ Treg in the MLNs (fig 8C). On the other hand, addition of mASCs to activated MLN cells isolated from DSS-treated mice inhibited their proliferative response and production of IFNγ in vitro, while IL10 secretion was stimulated (fig 8D). These findings suggest that mASCs suppress effector Th1 responses and induce T cells with suppressive functions.

Syngeneic and allogeneic murine adipose-derived mesenchymal stem cells (mASCs) protects against dextran sulfate sodium (DSS)-induced colitis. (A) Acute colitis was induced in C57Bl/6 mice by administration of 5% DSS from day 0 to day 7, and treated intraperitoneally with allogeneic (isolated from Balb/c mice) or syngeneic (isolated from C57Bl/6 mice) ASCs (106 cells/mouse) at day 2. Disease activity index and survival were monitored for different days. n = 5–8 mice/group. *p<0.001 vs DSS-treated mice. (B) Cytokine contents in colonic protein extracts isolated (day 6) from colitic mice untreated or treated with allogeneic ASCs were determined. n = 4 mice/group. *p<0.001 vs DSS-treated mice. (C) Lymphocytes from mesenteric lymph nodes (MLNs) isolated at day 6 in both groups were analysed for CD4 and CD25, and intracellular cytokine and FoxP3 expression by flow cytometry. Double staining for interferon γ (IFNγ)/interleukin 2 (IL2), IL4/IL10 and CD25/FoxP3 expression was performed in gated CD4 T cells. Numbers represent the percentage of cells in each quadrant. n = 4 mice/group. (D) MLN cells (105) isolated from DSS-treated mice at the peak of disease (day 6) were activated with phorbol 12-myristate 13-acetate +concanavalin A and co-cultured with or without allogeneic mASCs (2×104). The proliferative response was determined after 96 h culture, and cytokine contents in the supernatants were determined after 48 h. n = 4. *p<0.001 vs untreated MLN control samples. MIP-2, macrophage inflammatory protein-2; RANTES, regulated on activation normal T cell expressed and secreted; TNFα, tumour necrosis factor α.
hASCs protect against lethal endotoxemia
We finally investigated whether the therapeutic effect of hASCs could be extended to other inflammatory disorders caused by systemic infection with gut bacteria. We evaluated the action of hASCs in systemic inflammation and sepsis by using a model of endotoxaemia induced by a high dose of endotoxin and a model of diffuse peritonitis induced by caecal ligation and puncture (CLP), which mimics the clinical situation of patients with colonic leakage following surgical procedures.23 Treatment with hASCs or mASCs protected against mortality caused by endotoxin and CLP (fig 9A). The pathogenesis of sepsis is characterised by overwhelmed inflammatory and immune responses that can lead to tissue damage, multiple organ failure and death. hASC injection to septic animals decreased the levels of inflammatory mediators (TNFα, IL6, IL1β, IL12, IFNγ, RANTES and MIP-2) and increased IL10 in the major affected organs and diminished the inflammatory infiltration into the peritoneal cavity, lung, liver and intestine (fig 9B–E). These results indicate that hASCs rescue mice from endotoxemic death by downregulating the exacerbated inflammatory response characteristic of this disorder.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Human adipose-derived mesenchymal stem cells (hASCs) protect against lethal sepsis. Mice (10 mice/group) were injected intraperitoneally with lipopolysaccharide (LPS; 400 μg/mouse) and treated intraperitoneally with medium or 106 hASCs (3×105 when indicated) 30 min later. Alternatively, mice (8–10/group) were subjected to caecal ligation and puncture (CLP) and treated intraperitoneally with 106 hASCs or allogeneic murine ASCs (mASCs) 4 h later. (A) Survival was monitored every 12 h. (B) Cytokine and chemokine levels were determined by ELISA in protein extracts from liver, intestine and lung isolated 6 h after LPS injection. (C) Cytokine levels were determined in sera isolated 18 h after the CLP procedure. (D) The number of inflammatory cells infiltrating the peritoneal cavity was determined 6 h after LPS injection and 18 h after CLP. (E) Neutrophil infiltration in target organs was determined by measuring myeloperoxidase (MPO) activity in protein extracts obtained 6 h after LPS injection and 18 h after CLP. (F) Peritoneal macrophages (5×104) isolated from mice with CLP (n = 4) were cultured with medium (unstimulated) or with LPS (stimulated) in the absence (None) or presence of hASCs (5×104). When indicated, neutralising anti-interleukin 10 (IL10) antibodies were added to co-cultures. After 24 h, cytokine contents in the supernatants were determined by ELISA. (G) Peritoneal lavage, blood, spleen and liver were obtained 24 h after the CLP procedure (6 mice/group). Bacterial loads expressed as the number of colony-forming units (CFU) per ml were determined in the fluids and organ homogenates. *p<0.001 vs controls with LPS alone or CLP alone. ^p<0.001 vs hASC–macrophage co-cultures without anti-IL10 antibodies. IFNγ, interferon γ; MIP-2, macrophage inflammatory protein-2; RANTES, regulated on activation normal T cell expressed and secreted; TNFα, tumour necrosis factor α.
To investigate whether hASCs regulated the inflammatory response by acting directly on inflammatory cells, we co-cultured hASCs with activated macrophages isolated from septic mice. hASCs inhibited TNFα and IL12 production by macrophages, especially under inflammatory stimulation (fig 9F). This inhibitory effect was partially dependent on IL10 secretion (fig 9F). Accordingly, conditioned medium from activated hASC cultures significantly inhibited TNFα and IL12 production by macrophages (not shown).
Because CLP leads to a focal infection that subsequently becomes systemic, and persistence of the local bacterial infection plays a critical role in sepsis mortality, ASC treatment could influence the natural response against infection. Contrary to the expected results, ASC-treated septic mice had lower peritoneal bacterial counts than untreated septic mice (fig 9G), suggesting that ASCs could promote bactericidal activities by themselves or on other cell types. However, mASCs did not directly kill Escherichia coli in vitro in the absence of other cells (not shown).
DISCUSSION
Current therapy for CD is directed toward diminishing the inflammatory response and treating the sequelae of uncontrolled inflammation. To date, it has not been possible to prevent the disease or to arrest the disease process completely through medical treatment. In this study, we propose a novel cell-based therapeutic strategy for IBD by using adult MSCs isolated from adipose tissue. hASCs provided a highly effective treatment for both acute and chronic colitis induced by DSS. A single systemic injection of hASCs at the onset of the disease ameliorated the clinical and histopathological signs of the wasting disease, reducing the high mortality caused by this syndrome. The beneficial effect of ASCs on IBD was not restricted to a xenogeneic system. Both syngeneic and allogeneic mASCs were as efficient as hASCs in ameliorating the colitis. These findings suggest that the immunosuppressive action of ASCs is non-major histocompatibilty complex (MHC)-restricted and that the infused ASCs are immunotolerated by the host, which is very convenient for a future clinical application of these cells in IBD.
There are several potential mechanisms for the therapeutic effect of ASCs on the effector phase of colitis. Similar to CD, DSS-induced colitis is characterised by transmural inflammation of the colon owing to an IL12-driven, Th1 cell-mediated response to colonic and luminal antigens.1–3 Thus, innate and adaptive immune responses overlap during the effector phase of bowel inflammation, involving multiple inflammatory mediators. ASCs regulate the two arms of the immune response involved in the pathogenesis of IBD. ASCs strongly reduced colonic inflammation by downregulating the production of a wide panel of inflammatory/cytotoxic mediators by mucosal immune cells, and by increasing the levels of the anti-inflammatory cytokine IL10. Moreover, the inhibition of the production of chemokines is critical to explain the absence of inflammatory infiltrates in the lamina propria of ASC-treated mice.24 Deactivation of resident and infiltrating mucosal activated macrophages could be a major mechanism involved in the anti-inflammatory action of ASCs in colitis, and IL10 seems to play a critical role in this suppressive effect. ASCs only produce IL10 after cell contact with macrophages, and this contact seems to commit macrophages to produce high amounts of IL10. In agreement with our data, BM-MSCs stimulate the production of IL10 by monocytes and DCs.6 25 26
Interestingly, based on the expression of a particular set of chemokine receptors (ie, CCR1, CCR5 and CCR9), which have been involved in cell migration to inflamed tissues, hASCs rapidly and specifically target secondary lymphoid organs and the inflamed colon after their infusion. A recent study demonstrated that after the injection of syngeneic ASCs into the colonic mucosa of TNBS-colitic rats, ASCs preferentially localise in damaged/inflamed gut segments, where they accelerate the recovery of injured tissue by producing various regenerative factors.27 In our study, hASCs were detectable in the recipient for no more than 5 days postinjection, a time lag long enough to exert long-term immunosuppressive activities and to instruct other cells to turn down the exacerbated immune reaction locally and in lymphoid organs.
On the other hand, hASCs decreased the expression of Th1-type cytokines in the colon. The inhibition of Th1 responses might be the result of a direct action on cells of the draining MLNs because MLN cells obtained from hASC-treated animals were refractory in vitro to Th1 restimulation. In contrast, ASC treatment increased the production of IL10, but not IL4, in MLN CD4 T cells, arguing against a predominant Th2 response. IL10 is a signature cytokine for Treg, which play a key role in the control of self-antigen-reactive T cells in vivo.20–22 Besides inhibiting the inflammatory response, ASCs induce IL10-secreting Treg in colitic mice. These Treg could suppress effector T cells against colonic antigens. Indeed, ASC treatment of colitic mice increased the number of MLN Foxp3+CD4+CD25+ Treg. Moreover, CD4 cells isolated from hASC-treated mice showed profound beneficial effects on colitis. This is in agreement with recent works reporting the in vitro generation of Treg by human BM-MSCs,28–30 and that injection of allogeneic BM-MSCs11 or mASCs31 to arthritic mice prevented disease progression by inducing the emergence of antigen-specific Treg. The involvement of Treg and IL10 in the therapeutic effect of ASCs on colitis is supported by the fact that in vivo depletion of IL10 and CD25+ T cells partially reversed their protective actions.
This work also provides the important evidence that ASCs might represent feasible therapeutic agents to treat sepsis. Sepsis, a life-threatening complication of bacterial infections and the most common cause of death in intensive care units, is characterised by a hyperactive and out-of-balance network of endogenous proinflammatory cytokines. Most therapeutic strategies have targeted proinflammatory mediators, but they did not improve survival of patients when studied in large multicentre clinical trials.32 Treatments designed to block one single cytokine, such as TNFα and IL1β, have shown limited efficacy probably due to their early and transient kinetics. Here, we show that ASCs protect against the mortality in two models of severe endotoxaemia and sepsis. hASCs function at several levels to regulate many crucial aspects of sepsis, including reduction of the systemic levels of a wide panel of inflammatory cytokines and chemokines and inhibition of leucocyte infiltration of various targets organs. Once again, because macrophages and monocytes play a central role in the production of inflammatory mediators during sepsis, they appear as major cell targets in the protective effect of ASCs in sepsis. Importantly, a recent work demonstrated that hASCs express different Toll-like receptors and become activated in response to different bacterial products.33
Taken together, our data demonstrate the capacity of hASCs to suppress acute inflammatory autoimmune responses in mice by inhibiting the inflammatory and Th1 responses. This might offer a therapeutic advantage over neutralising antibodies directed against a single mediator. In addition, the maintenance of tolerance by IL10-secreting Treg could be related to the resistance to disease recurrence observed in hASC-treated mice, mainly because they persisted for a long period of time. We envisage that once the benefits and risks associated with the injection of hASCs are well defined, the use of hASCs will open up new perspectives for a cell-based treatment of IBD.
REFERENCES
Supplementary materials
Supplementary Data
This web only file has been produced by the BMJ Publishing Group from an electronic file supplied by the author(s) and has not been edited for content.
- Data supplement 1 - Online supplement
Footnotes
See Commentary, p 898
▸ Competing interests: Declared (the declaration can be viewed on the Gut website at http://www.gut.bmj.com/supplemental). Additional methods, figures and a table are published online only at http://gut.bmj.com/content/vol58/issue7
Funding: This work was supported by Ministry of Health, Ministry of Education and Science (PETRI), RETICS, Cellerix SA and Junta de Andalucia. DB had been the recipient of a Marie Curie International Integration Grant.