Article Text

Abstract
Background Neutrophils are accumulated in inflamed mucosa of IBD and play an important role in the pathogenesis. CD177 is expressed in neutrophils specifically and upregulated during inflammation. However, the role of CD177+ neutrophils in pathogenesis of IBD remains elusive.
Materials and methods Expression of CD177 was analysed in peripheral blood and intestinal mucosa from patients with IBD using quantitative RT-PCR, flow cytometry and immunohistochemistry. CD177+ and CD177− neutrophils were isolated to determine gene differences by RNA sequencing. Colitis was established in CD177−/− and wild-type mice in response to dextran sulfate sodium (DSS) insults to determine the role of CD177+ neutrophils in IBD.
Results CD177+ neutrophils were markedly increased in peripheral blood and inflamed mucosa from patients with active IBD compared with healthy controls. RNA sequencing revealed that differential gene expression between CD177+ and CD177− neutrophils from patients with IBD was associated with response to bacterial defence, hydrogen peroxide and reactive oxygen species (ROS). CD177+ neutrophils produced lower levels of proinflammatory cytokines (ie, interferon-γ, interleukin (IL)-6, IL-17A), but higher levels of IL-22 and transforming growth factor-β, and exhibited increased bactericidal activities (ie, ROS, antimicrobial peptides, neutrophil extracellular trap) compared with CD177− subset. CD177−/− mice developed more severe colitis on DSS insults compared with wild-type mice. Moreover, CD177 deficiency led to compromised intestinal barrier and impaired antibacterial immunity through decreased production of IL-22 by CD177− neutrophils.
Conclusions CD177+ neutrophils represent functionally activated population and play a protective role in IBD through increased bactericidal activity and IL-22 production. Targeting CD177+ neutrophils may be beneficial for treatment of IBD.
- IBD BASIC RESEARCH
- INTERLEUKINS
- MUCOSAL IMMUNOLOGY
- CELLULAR IMMUNOLOGY
Statistics from Altmetric.com
Significance of this study
What is already known on this subject?
Neutrophils play an indispensable role in the pathogenesis of IBD.
CD177 is expressed in neutrophils specifically and involved in the pathogenesis of ANCA-associated systemic vasculitis and systemic lupus erythematosus.
What are the new findings?
CD177 is highly expressed in peripheral neutrophils and inflamed mucosa from patients with IBD.
Differential gene expression between CD177+ and CD177− neutrophils from patients with IBD is associated with response to bacterial defence, hydrogen peroxide and reactive oxygen species.
CD177+ neutrophils produce low levels of proinflammatory cytokines, but high levels of interleukin (IL)-22, and exhibit increased bactericidal activities.
CD177 depletion aggravates dextran sulfate sodium-induced acute colitis.
CD177+ neutrophils-derived IL-22 promotes intestinal barrier function and enhances antibacterial immunity.
How might it impact on clinical practice in the foreseeable future?
CD177+ neutrophils play a protective role in the pathogenesis of IBD through increased bactericidal activity and IL-22 production. Targeting CD177+ neutrophils may serve as a novel therapeutic approach in the management of human IBD.
Introduction
IBD, comprising Crohn's disease (CD) and ulcerative colitis (UC), are chronic remittent and progressive inflammatory disorders that affect the GI tracts and may be associated with an increased risk for colon cancer.1 ,2 Although their exact aetiology and pathology are still not clear, IBD appears to be considered as the result from abnormal immune response to intestinal microbiota in genetically susceptible individuals.3–6 The hallmarks of IBD are dysregulated intestinal immune responses,7–10 in which neutrophils are accumulated in inflamed mucosa at the early stage of inflammatory response and play an indispensable role in the pathogenesis. Neutrophils are short-lived effector cells in innate immune system, which are the most abundant leucocyte population in the blood and also typically the first leucocytes to be recruited into the inflammatory areas.11 ,12
Under inflammatory conditions, bacteria-derived lipopolysaccharide (LPS), formyl-methionylleucyl-phenylalanine, DNA or inflammatory mediators, such as interleukin (IL)-8, chemokine (C-X-C motif) ligand 1 (CXCL-1), growth related oncogene-α, C5a and leukotriene B4, induce neutrophil mobilisation, including exiting the circulation, migrating across the lamina propria (LP) and finally crossing the epithelial barrier.12 When contacting with invading microbes, neutrophils are adapted to perform a wide variety of antimicrobial functions, such as phagocytosis, release of reactive oxygen species (ROS), degranulation and formation of neutrophil extracellular traps (NET) to eliminate intestinal microbial infection.12 ,13 Recently, we have demonstrated in a microbiota antigen-specific T cell-dependent chronic model that neutrophils protect the intestines from chronic inflammation through production of IL-22.14 In acute murine colitis induced by dextran sulfate sodium (DSS), colon-infiltrating neutrophils produce high levels of IL-22 and target colonic epithelial cells to upregulate the antimicrobial peptides, which further contribute to the resolution of colonic inflammation.15 Moreover, by adhesion to intercellular adhesion molecule-1, neutrophils result in the activation of Akt and β-catenin signalling pathways, leading to epithelial proliferation and mucosal wound healing.16 However, it is still unclear which subsets of neutrophils play a protective role in intestinal mucosal inflammation, particularly in IBD.
CD177, also known as human neutrophil antigen NB1 or polycythaemia rubra vera-1, is an approximately 58−64 kDa glycosylphosphatidylinositol-anchored glycoprotein expressed exclusively in neutrophils, neutrophilic myelocytes and metamyelocytes.17 ,18 It is expressed in 45%−60% of peripheral neutrophils in healthy donors, but upregulated during acute bacterial infections or following granulocyte colony-stimulating factor treatment.19 It has been demonstrated that CD177 in neutrophils can bind platelet endothelial cell adhesion molecule-1 (or CD31), mediate neutrophil-endothelial cell interaction and promote neutrophil transmigration.18 Moreover, CD177 is associated with expression of granule protein and involved in the pathogenesis of antineutrophil cytoplasmic antibody (ANCA)-associated systemic vasculitis and systemic lupus erythematosus (SLE).19 ,20 However, it is still unclear whether CD177+ cells are a specific functional subset of neutrophils to regulate the pathogenesis of IBD.
In the current study, we demonstrated that CD177+ neutrophils were upregulated in both peripheral blood neutrophils and inflamed intestinal mucosa of patients with IBD. CD177+ neutrophils produced less proinflammatory cytokines but higher levels of IL-22, and exhibited increased bactericidal activities compared with CD177− subset. Furthermore, CD177−/− mice developed more severe colitis on DSS insults. The diversity and abundance of gut bacterial microbiota were decreased in CD177−/− mice with colitis compared with controls. CD177 deficiency led to compromised intestinal barrier function and impaired antibacterial immunity through decreased production of IL-22 by CD177− neutrophils. Collectively, our data indicate that CD177+ neutrophils represent a functional activated subset of neutrophils, which protect the intestines from inflammation.
Materials and methods
Patients
All patients with IBD were recruited from the Department of Gastroenterology, the Shanghai Tenth People's Hospital of Tongji University (Shanghai, China) from February 2014 to October 2016. EDTA anticoagulated blood samples (20 mL) were obtained from patients with active CD (A-CD, n=50), patients with CD in remission (R-CD, n=38), patients with active UC (A-UC, n=44), patients with UC in remission (R-UC, n=35) and healthy controls (HC, n=37) after overnight fasting. Colon biopsy samples were obtained from A-CD (n=47), R-CD (n=36), A-UC (n=40), R-UC (n=32) and HC (n=33) who underwent endoscopy. The diagnoses for CD or UC were based on clinical characteristics, radiological and endoscopic examination and histological findings. The clinical characteristics of these patients are shown in online supplementary table S1. International standard criteria such as Crohn's Disease Activity Index (CDAI) and Mayo scores were used to assess the severity of disease in patients with CD or UC, respectively.21 This study was approved by the Institutional Review Board for Clinical Research of the Shanghai Tenth People's Hospital of Tongji University. Written informed consent was also obtained from all subjects before study.
supplementary table
Isolation of CD177+ and CD177− neutrophils
EDTA anticoagulated blood samples (20 mL) were obtained from patients with IBD and HC as described above.21 After gradient centrifugation and red blood cell lysis, neutrophils were obtained. CD177+ neutrophils were isolated by using antihuman CD177-magnetic sorting (Miltenyi Biotec; Gladbach, Germany), and CD177− neutrophils were then isolated by using antihuman CD66b-magnetic sorting (Miltenyi Biotec). The purity of CD177+ and CD177− neutrophils was examined on a BD FACSCanto II (BD Biosciences; San Diego, California, USA), and >95% was used for further experiments.
Analysis of NET formation by fluorescence microscopy
CD177+ and CD177− neutrophils were isolated from peripheral blood and stimulated with LPS (200 ng/mL) for 2 hours. They (2×105) were seeded on the poly-l-lysine-coated glass slides, fixed with 4% paraformaldehyde, and then stained with Hoechst 33342 (1:1000). NETs were visualised on a confocal microscopy (LSM 710, ZEISS; Oberkochen, Germany).
Mice
C57BL/6 CBir1-specific TCR transgenic (CBir1 TCR Tg) mice were bred and maintained in the Animal Facilities at University of Texas Medical Branch.22 ,23 C57BL/6 Rag-1−/− mice were purchased from the Nanjing Biomedical Research Institution of Nanjing University (Nanjing, China) and bred at the animal facility of the Shanghai Tenth People's Hospital, Tongji University. C57BL/6 CD177−/− mice were purchased from the KOMP in University of California (Davis, California, USA) and bred at the animal facility of the Shanghai Tenth People's Hospital. These mice and C57BL/6 wild-type (WT) mice were raised in microisolator cages with filtered air under specific pathogen-free conditions, and free access to sterile water and autoclaved food. Mice used in the experiment were aged 8–10 weeks and 20−25 g of weight. Animal studies were reviewed and approved by the Institutional Animal Care and Use Committee of the UTMB and the Tongji University, respectively.
DSS-induced colitis model in mice
DSS-induced colitis model was established using a method described previously.24 ,25 Briefly, WT and CD177−/− mice (n=12 per group) were given 2.0% DSS (molecular mass, 36 000–50 000, MP Biomedicals, Solon, Ohio, USA) in the drinking water for continuous 7 days, then followed with regular water for another 3 days. Characteristics of acute colitis were observed daily, including diarrhoea, bloody stools, body weight and survival rates. The histological grading of colonic inflammation was graded from 0 to 4 as reported previously.21
Isolation of intestinal epithelial cells and lamina propria mononuclear cells
As described previously,22 ,26 the colon was removed from mice, sliced into 0.5−1.0 cm pieces and washed with cold phosphate-buffered saline (PBS) to remove faecal contents. After the digestion by EDTA, primary intestinal epithelial cells (IECs) were collected in the supernatants, and further purified via density gradient centrifugation with 20% and 40% Percoll-Roswell Park Memorial Institute (RPMI) solution. Colon tissues were then digested by collagenase A (Sigma-Aldrich, St. Louis, Missouri, USA), and single cell suspension was collected and further purified via density gradient centrifugation with 40% and 70% Percoll-RPMI solution. Lamina propria mononuclear cells (LPMCs) were collected from the interface and suspended in 10% fetal bovine serum (FBS)-RPMI medium.
Coculture of epithelial cells and neutrophils
Neutrophils were isolated from the bone marrow of WT and CD177−/− mice, and stimulated with LPS (200 ng/mL) for 2 hours. Colon-derived MC38 epithelial cells were cultured in RPMI-1640 medium supplemented with 10% FBS, penicillin (100 U/mL), streptomycin (100 μg/mL) at 37°C with 5% CO2. MC38 cells (1×105) were seeded into 12-well culture plates and cultures in RPMI-1640 medium. Forty-eight hours later, WT or CD177−/− neutrophils were added in the medium at the effector-to-target cell ratio of 5:1 in the presence of IL-23 (10 ng/mL). After 24 hours, MC38 cells were washed with PBS, and processed for RNA isolation.
Statistical analysis
All data were expressed as mean±SEM, and analysed using SPSS V.20.0 (SPSS, Chicago, Illinois, USA). Statistical comparisons were performed using an unpaired two-tailed Student's t-test. Pearson's correlation was performed to analyse the correlation of CD177 expression in intestinal mucosa with CDAI and Mayo index; *p<0.05, **p<0.01 and ***p<0.001 were considered to be statistically significant.
See online supplementary materials and methods for additional information.
supplementary methods
Results
CD177 expression and CD177+ neutrophils are increased in mice with chronic colitis and patients with active IBD
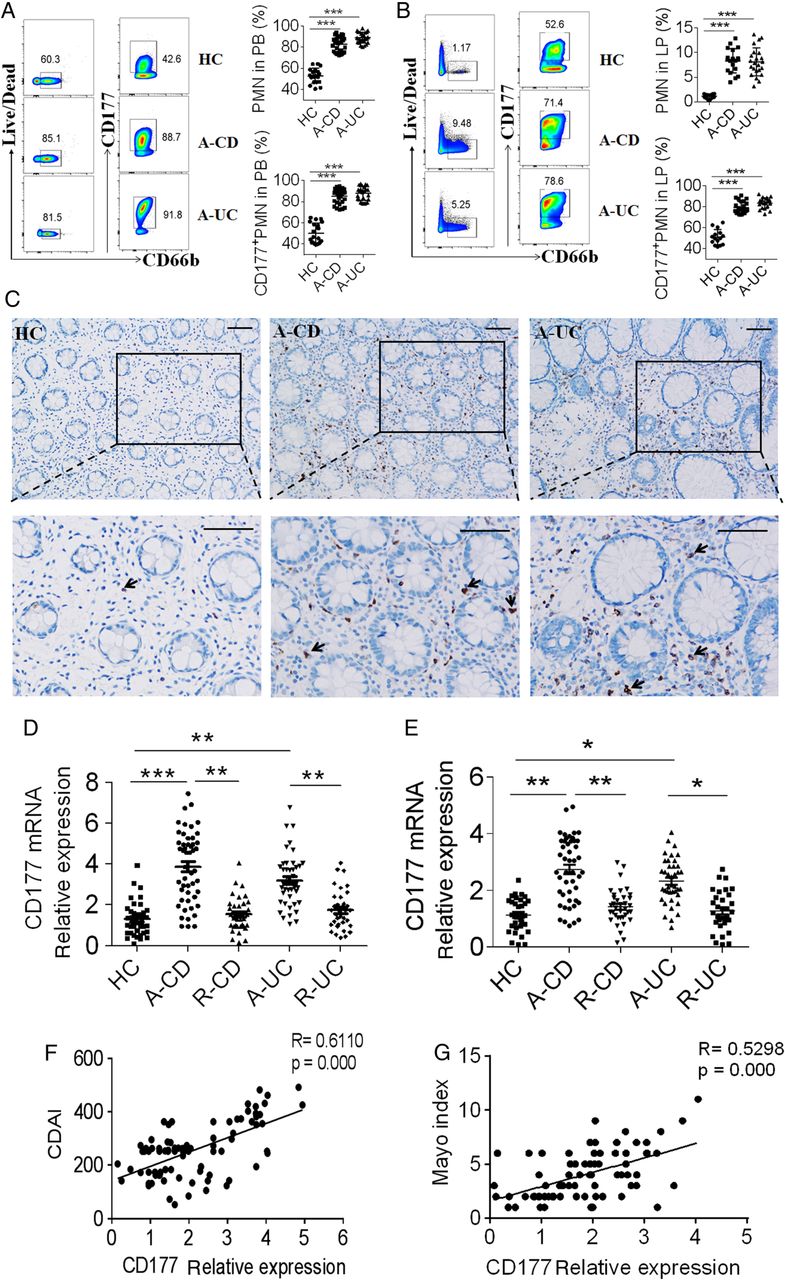
CD177 has been shown to be exclusively expressed in neutrophils. To determine the potential role of CD177+ neutrophils in intestinal inflammation, we determined kinetic expression of CD177 in intestines in a microbiota antigen-specific T cell-mediated chronic colitis model, in which CBir1 TCR-CD4+ T cells, which are specific for an immunodominant microbiota antigen CBirl flagellin, were transferred into Rag-1−/− mice, in which the recipient mice develop mild intestinal inflammation starting from week 2, and demonstrate severe colitis at week 4 after T cell transfer.22 Neutrophils accumulated in the LP of colon from week 1 and peaked at week 2 after CD4+ T cell transfer, while CD4+ T cells were increased continually during the development of colitis (see online supplementary figure S1A). Interestingly, the same pattern of CD177 mRNA was also observed in the bone marrow cells and colonic tissues (see online supplementary figure S1B), indicating that neutrophils and CD177 may participate in early stage of intestinal mucosal inflammation. We also analysed CD177+CD66b+ neutrophils from patients with IBD by flow cytometry, and found that they were significantly increased in peripheral blood and inflamed colonic mucosa from patients with active IBD compared with HC (figure 1A,B). To localise CD177+ neutrophils in inflamed mucosa of IBD, colon sections from patients with active IBD and HC were stained for CD177 by immunohistochemistry. CD177+ neutrophils were markedly increased in inflamed mucosa from patients with active IBD compared with HC (figure 1C). Moreover, the levels of CD177 mRNA were also increased in both peripheral neutrophils (figure 1D) and inflamed mucosa (figure 1E) from patients with IBD compared with that in healthy donors, whereas no statistical difference was observed between CD and UC groups. Since the CDAI and Mayo index are routinely used to evaluate the disease severity of CD and UC, respectively,21 ,27 we then analysed the correlation of CD177 expression with the CDAI and Mayo index, respectively. Interestingly, we observed that CD177 expression in inflamed mucosa from patients with CD or UC was positively correlated with the CDAI and Mayo index, respectively (figure 1F,G).
supplementary figures

CD177 is highly expressed in patients with IBD. The percentages of neutrophils and CD177+ neutrophils in peripheral blood (A) and colon biopsies (B) were analysed by flow cytometry. Neutrophils were isolated from peripheral blood of healthy controls (HC, n=20), patients with active Crohn's disease (A-CD, n=29) and patients with active UC (A-UC, n=26). Lamina propria mononuclear cells (LPMC) were isolated from biopsies of colonic tissues of HC (n=15), A-CD (n=23) and A-UC (n=20). CD66b+ neutrophils and CD177+CD66b+ neutrophils were analysed by flow cytometry. (C) Representative images of immunohistochemical staining of CD177 expression in inflamed colon from patients with A-CD, A-UC and normal colonic mucosa from HC. The arrows indicate CD177+ cells. Scar bar represents 50 μm. (D) Expression of CD177 mRNA in peripheral neutrophils of patients with IBD. Peripheral neutrophils were isolated from patients with A-CD (n=50), A-UC (n=44), patients with CD in remission (R-CD, n=38), patients with UC in remission (R-UC, n=35) and HC (n=37). CD177 mRNA was examined by quantitative RT-PCR (qRT-PCR). (E) Expression of CD177 mRNA in inflamed mucosa of patients with IBD. RNA was isolated from colon biopsies from HC (n=33), and patients with A-CD (n=47), A-UC (n=40), R-CD (n=36) and R-UC (n=32). CD177 mRNA was then analysed by qRT-PCR. Gene expression was normalised to GAPDH in each group. *p<0.05, **p<0.01, ***p<0.001. Data are pooled from three independent experiments. (F) The correlation analysis was performed between Crohn's Disease Activity Index (CDAI) and CD177 expression in inflamed mucosa from patients with CD (R=0.6110, p=0.000). (G) The correlation analysis was performed between the Mayo index and CD177 expression in inflamed mucosa from patients with UC (R=0.5298, p=0.000). LP, lamina propria; PB, peripheral blood; PMN, polymorphonuclear leukocytes.
CD177+ neutrophils demonstrate enhanced bactericidal activity with producing high levels of ROS, MPO, NET and antimicrobial peptides
To determine whether there was a difference in morphology between CD177+ and CD177− neutrophils, we isolated CD177+ and CD177− neutrophils from peripheral blood of healthy donors using immunomagnetic beads. The purity of CD177+ and CD177− neutrophils used in the experiment was usually >95% as assessed by flow cytometry (see online supplementary figure S2A). Cell smear of highly purified CD177+ and CD177− neutrophils was stained by Wright-Giemsa. As shown in online supplementary figure S2B, no morphological difference was observed between these two cell subsets. Moreover, no significant difference was found in granules presented in the cytoplasm between CD177+ and CD177− neutrophils under transmission electron microscopic images (see online supplementary figure S2C). We then investigated if there were differences in gene expression profiles between CD177+ and CD177− neutrophils. We isolated CD177+ and CD177− neutrophils from patients with CD, UC and healthy donors, and performed RNA sequencing. Significant gene expression differences were observed between CD177+ and CD177− subsets, especially from patients with CD or UC (see online supplementary figure S3A–-C). Pathway analysis of differential gene profiles revealed that the differential gene expression was associated with defence response, response to bacteria and wounding, inflammatory response, response to ROS and hydrogen peroxide in patients with CD or UC (figure 2B,C), whereas in healthy donors the gene differences were mainly associated with carbohydrate biosynthetic process, lipid biosynthetic process and glycoprotein metabolic process (figure 2A). We next compared differentially expressed genes in CD177+ neutrophils between patients with IBD and HC. Pathway analysis of differential gene profiles revealed that differentially expressed genes in CD177+ neutrophils between patients with IBD and HC were associated with defence response and inflammatory immune response (see online supplementary figure S4A–C).
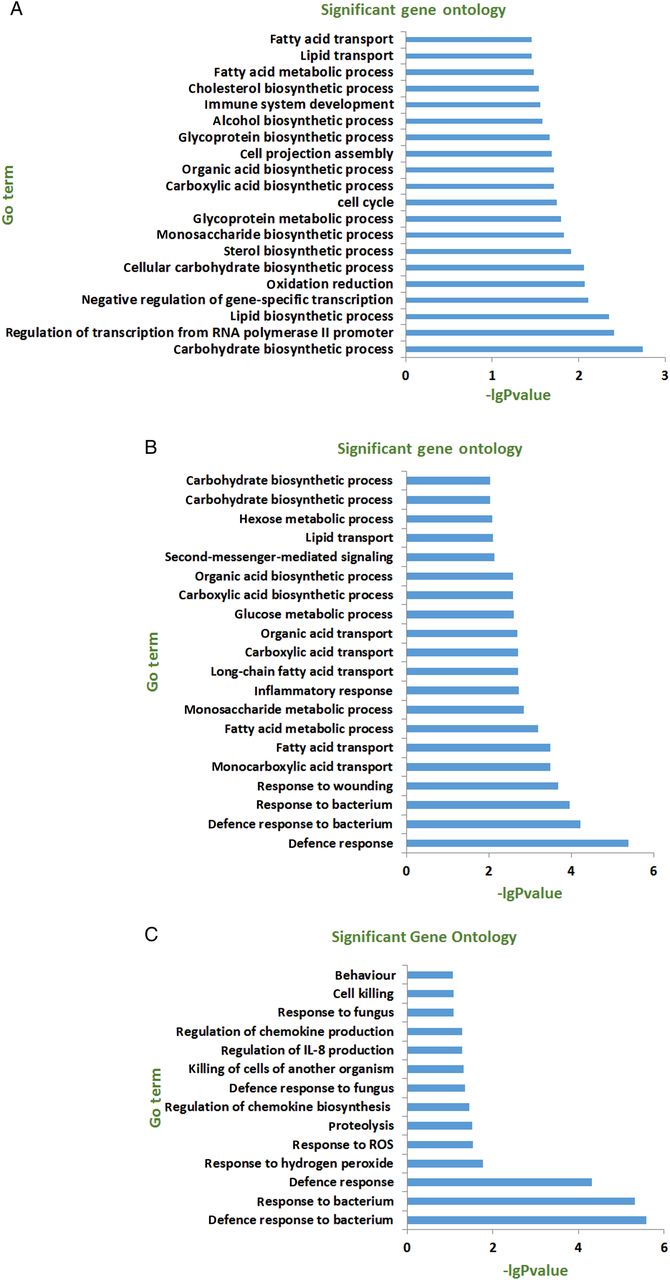
Differential gene profiles in CD177+ and CD177− neutrophils. Highly purified CD177+ and CD177− neutrophils were isolated from peripheral blood of patients with IBD and healthy donors using immunomagnetic beads, and used to detect transcriptome differences by RNA sequencing. Pathway analysis of differential gene expression (fold change >2.0) in healthy controls (HC) (A, n=4) and patients with patients with active Crohn's disease (A-CD) (B, n=3) and patients with active UC (A-UC) (C, n=3). IL, interleukin; ROS, reactive oxygen species.
Neutrophils have long been viewed as the effector cells with the primary role in resistance to the invading pathogens, and are classically characterised by the ability to phagocytose microorganisms, release antibacterial proteins (cathepsins, defensins, lactoferrin, lysozyme) from their granules, and produce ROS.11 ,28 ,29 ROS are produced by the phagocyte NADPH oxidase and have important antimicrobial properties, which are crucial for host defence against microbial infections.30 RNA sequencing revealed that the functional differences between CD177+ and CD177− neutrophils were mainly reflected in defence response, response to ROS and hydrogen peroxide in patients with CD or UC (figure 2B,C), suggesting that CD177+ neutrophils play an important role in inflammatory response and defence response to invading microbiota. To further determine whether there was difference in antimicrobial activities between CD177+ and CD177− subsets, we measured ROS production by peripheral neutrophils under spontaneous and phorbol 12-myristate 13-acetate (PMA)-stimulated conditions using the Amplex Red assay, and found that relatively low levels of ROS and myeloperoxidase (MPO) were produced in untreated CD177+ and CD177− neutrophils. However, PMA-triggered CD177+ neutrophils produced significantly higher levels of ROS and MPO compared with CD177− neutrophils (figure 3A,B). Moreover, expression of S100A8, S100A9 and MPO was significantly higher in CD177+ neutrophils than those in CD177− subset (figure 3C–E). These data indicate that CD177+ neutrophils have enhanced bactericidal activities.

CD177+ neutrophils release more reactive oxygen species (ROS), MPO, neutrophil extracellular traps (NET) and antimicrobial peptides. The levels of ROS (A) and MPO (B) produced by peripheral neutrophils (1×104) from patients with active Crohn's disease (A-CD, n=6), patients with active UC (A-UC, n=6) and healthy controls (HC, n=6) were measured using Amplex Red Hydrogen Peroxide Assay Kit according to the manufacturer's instructions. *p<0.05, **p<0.01, ***p<0.001. (C−E) CD177+ and CD177− neutrophils isolated from peripheral blood of HC (n=9), and patients with A-CD (n=16) or A-UC (n=14) were stimulated with lipopolysaccharide (LPS) (200 ng/mL) for 2 hours. Expression of MPO (C), S100A8 (D) and S100A9 (E) was then analysed by quantitative RT-PCR and normalised to GAPDH. *p<0.05, **p<0.01, ***p<0.001. (F) CD177+ and CD177− neutrophils isolated from peripheral blood of HC (n=5), patients with A-CD (n=8) and patients with A-UC (n=8) were stimulated with LPS (200 ng/mL) for 2 hours and stained with Hoechst 33342. The formation of NET was examined by immunofluorescence microscopy (×200). Data are representative of three independent experiments. PMN, polymorphonuclear leukocytes.
Neutrophils can generate NET to trap and kill invading microorganisms.31 We then determined the formation of NET in neutrophils. CD177+ and CD177− neutrophils were isolated from peripheral blood of patients with IBD and healthy donors, stimulated in vitro with LPS and stained with Hoechst 33342, a dye that stains the primary component of NET and nucleic acid. NET formation was observed to be increased in LPS-stimulated CD177+ neutrophils compared with that in CD177− neutrophils. Moreover, NET formation was also increased in LPS-stimulated CD177+ neutrophils from patients with active IBD compared with that from HC. In contrast, NET formation was much less in unstimulated CD177+ and CD177− subsets (figure 3F). Collectively, these data indicate that CD177+ neutrophils increase bactericidal activities by releasing high levels of ROS, MPO, NET and antimicrobial peptides, which may protect the intestines from inflammation.
CD177+ neutrophils produce low levels of proinflammatory cytokines but high levels of IL-22
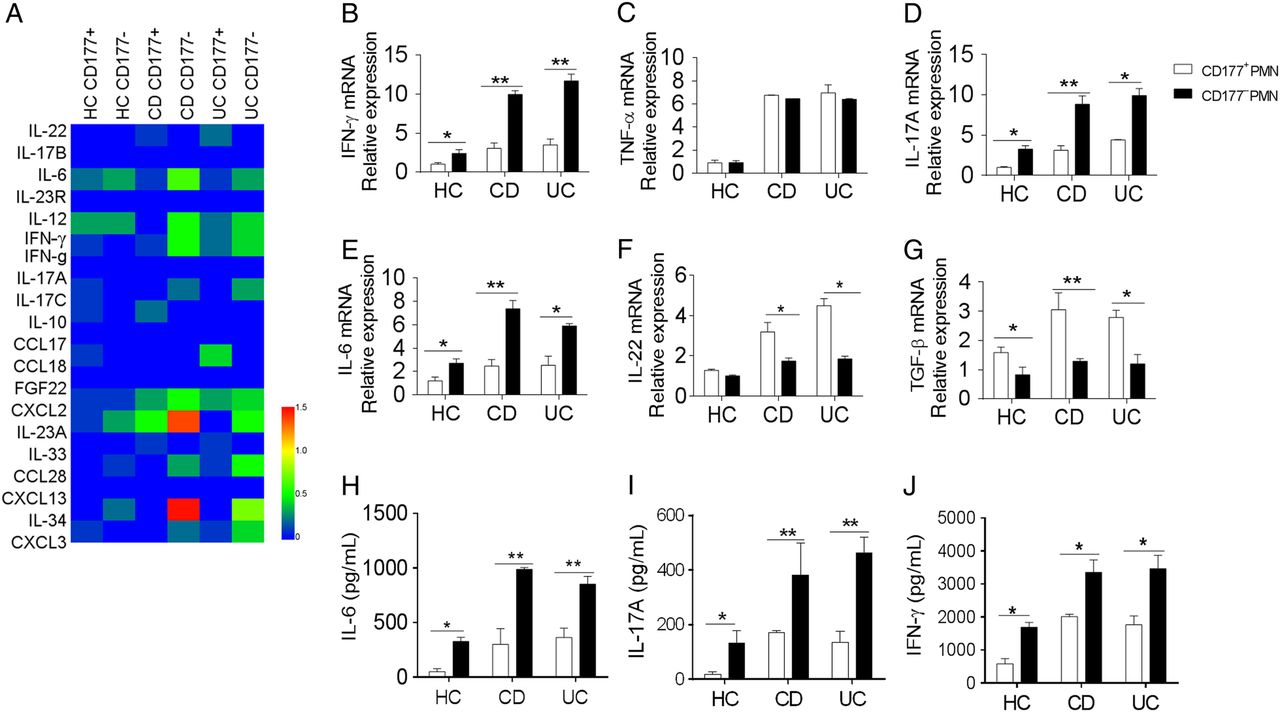
RNA sequencing also revealed CD177+ neutrophils produced low levels of proinflammatory cytokines (eg, IL-6, IL-12, IL-23 and interferon (IFN)-γ) and chemokines (eg, CXCL2, CCL28) compared with CD177− neutrophils, whereas CD177+ neutrophils produced more IL-22 than CD177− subset (figure 4A). To further confirm these data, we isolated CD177+ and CD177− neutrophils from peripheral blood of patients with active IBD and HC, and stimulated with LPS as indicated. As shown in figure 4B-J, CD177+ neutrophils produced higher levels of IL-22 and transforming growth factor (TGF)-β, but lower levels of IL-6, IL-17A and IFN-γ than CD177− neutrophils.
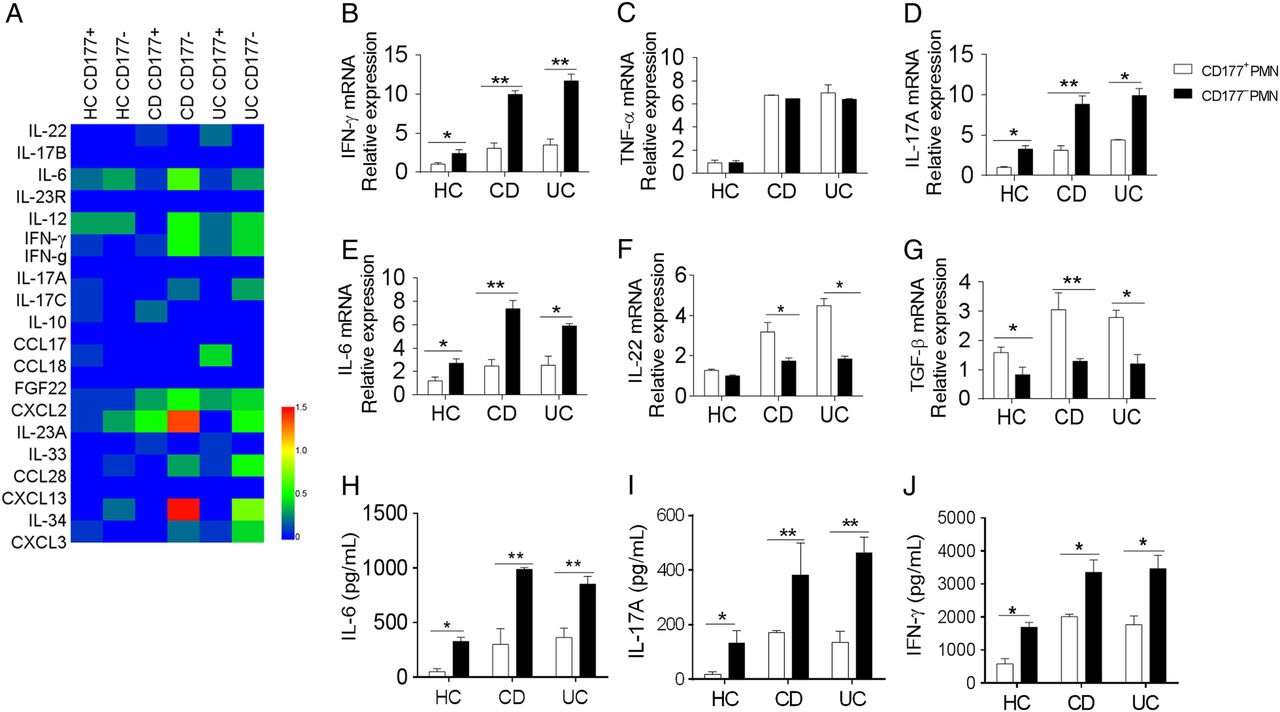
CD177+ neutrophils produce low levels of proinflammatory cytokines, but high levels of interleukin (IL)-22. (A) Heat maps of some differential genes in CD177+ and CD177− neutrophils by RNA sequencing. CD177+ and CD177− neutrophils were isolated from peripheral blood of healthy controls (HC, n=9), patients with Crohn's disease (CD, n=16) and patients with UC (n=14), and stimulated in vitro with lipopolysaccharide (LPS) (200 ng/mL) for 2 hours. Cytokines were analysed by quantitative RT-PCR and normalised to GAPDH (B–G). CD177+ and CD177− neutrophils (2×106) were isolated from peripheral blood of HC (n=6), patients with CD (n=10) and patients with UC (n=12). After pretreatment in vitro with LPS (200 ng/mL) for 2 hours, they (2×106) were then cultured in culture medium alone for further 24 hours, and supernatants were collected for the detection of IL-6 (H), IL-17A (I) and interferon (IFN)-γ (J) by ELISA. *p<0.05, **p<0.01. Representative results are shown from three independent experiments. TGF, transforming growth factor; TNF, tumour necrosis factor.
CD177−/− mice develop more severe colitis on DSS insults
To determine the role of CD177+ neutrophils in the pathogenesis of IBD, DSS-induced colitis model was used in WT and CD177−/− mice. As shown in figure 5A, the survival rate was significantly decreased in CD177−/− mice compared with WT mice during a period of 10-day observation. CD177−/− mice developed more severe colitis, characterised by a significant loss of body weight, more shorted colon length, higher levels of pathological scores compared with those in WT mice after DSS exposure (figure 5B–F). Colon tissues were then obtained from DSS-induced colitis mice, and total RNA was extracted to detect inflammatory cytokine expression. Expression of tumour necrosis factor (TNF)-α, IFN-γ, T-bet, IL-17A, IL-6, RORγt and IL-1β was significantly increased in CD177−/− mice, whereas IL-22 expression was decreased in CD177−/− mice. No significant expression of IL-4 and GATA3 was observed (see online supplementary figure S5). Moreover, fresh colon tissues were also obtained and cultured in vitro for 24 hours, the supernatants were collected to detect cytokine production by ELISA. We found that IL-17A, IFN-γ and IL-1β production were significantly higher in CD177−/− mice than those in WT mice, whereas IL-22 production was lower in CD177−/− mice, which was consistent with the results by quantitative RT-PCR (qRT-PCR) (see online supplementary figure S6). We then examined phenotypic profiles of T and B cells in LPMCs from WT and CD177−/− mice, and found that there were no differences of CD4+ T cell, CD8+ T and B cell population between WT and CD177−/− mice with or without DSS insult (see online supplementary figure S7). Gut microbiota has been implicated in the pathogenesis of IBD and microbial diversity is decreased in IBD.32 We collected faeces from CD177−/− mice and cohoused WT controls to assess microbial differences by high-throughput 16S ribosomal DNA gene sequencing. Shannon diversity index further proved more decreased microbial diversity in CD177−/− mice than that in WT mice (figure 5G), consistent with the severity of intestinal inflammation. Rank-abundance curve also demonstrated that species abundance was lower and species distribution was uneven in CD177−/− mice (see online supplementary figure S8A). Principal coordinates analysis (PCoA) on the basis of microbial composition revealed major differences between WT and CD177−/− mice after DSS exposure (see online supplementary figure S8B,C). The faecal microbiota in CD177−/− mice after DSS exposure was dominated by pathogenic bacteria (eg, Mucispirillum, Enterococcus, Turicibacter and Escherichia-Shigella), while there was a marked decrease of bacteria in Firmicutes and Bacteroidetes (figure 5H).

CD177 deficiency aggravates dextran sulfate sodium (DSS)-induced colitis in mice. Wild-type (WT) and CD177−/− mice (n=12 in each group) were given 2.0% DSS in drinking water for continuous 7 days, then followed with regular water for another 3 days. On day 10, all mice were sacrificed. (A) The survival rate in both groups after DSS insult. *p<0.05 vs DSS-exposed WT mice. (B) The changes of body weight were expressed as a percentage of the original weight at the start of the experiments during a period of 10-day observation. *p<0.05, **p<0.01 vs DSS-exposed WT mice at the same time points. (C) The length of colons was shown in the chart. **p<0.01, ***p<0.001. (D) Gross morphology of the colon on day 10 after DSS induction. (E) Representative H&E staining images of ascending colon (original magnification ×200). (F) The changes of pathological scores from colonic sections were calculated as indicated. **p<0.01. The faeces were collected from WT and CD177−/− mice with or without DSS-induced colitis, and used to detect the diversity of microbial communities by high-throughput 16S ribosomal DNA (rDNA) gene sequencing. (G) 16S rDNA Shannon diversity index in the faecal samples from WT and CD177−/− mice. (H) Heat map of bacterial genes of WT and CD177−/− mice. Data are representative of three independent experiments.
Mouse CD177+ neutrophils produce high levels of IL-22 and maintain epithelial barrier function
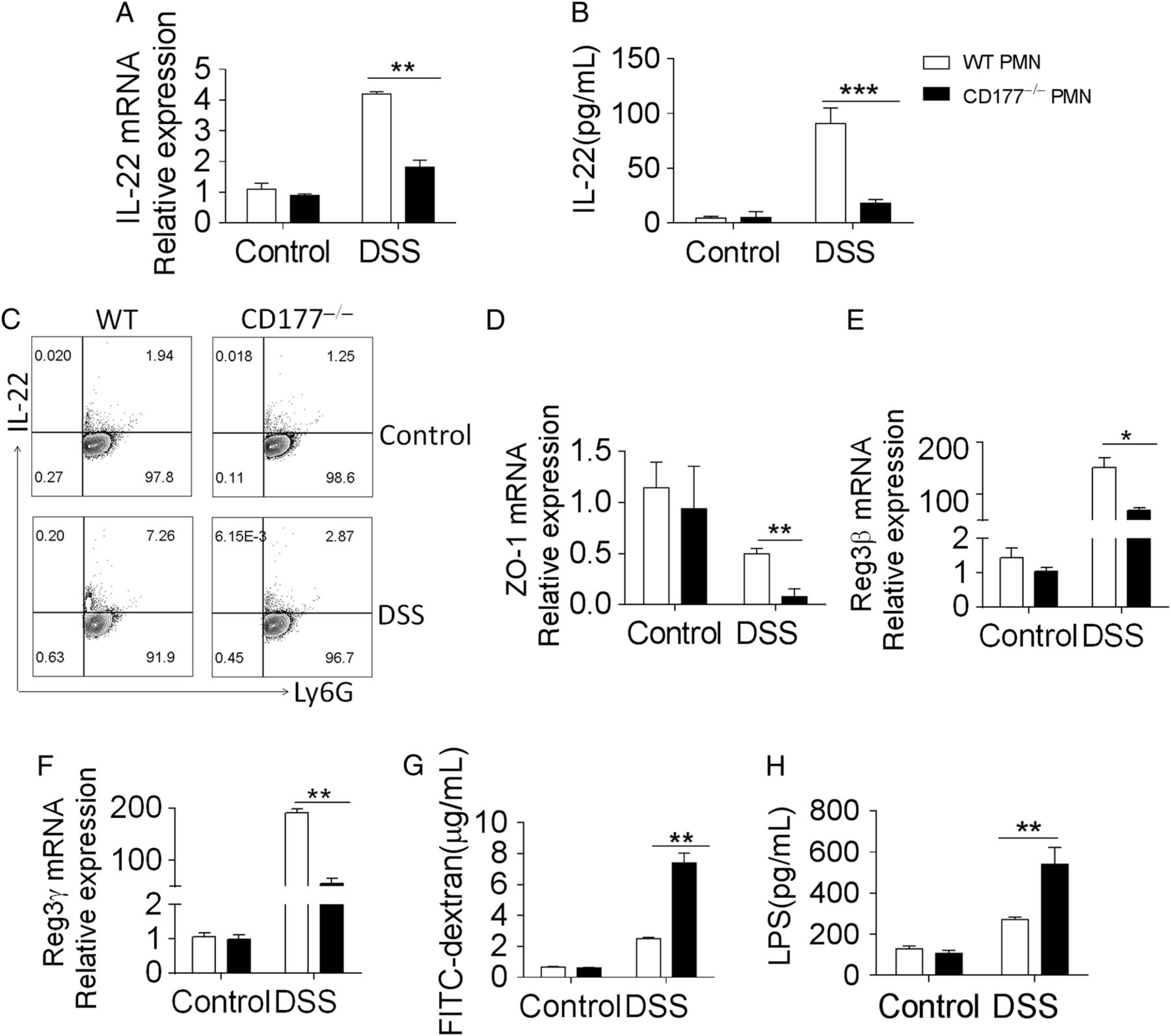
IL-22, belonging to IL-10 cytokine family, has been reported to be upregulated in IBD33 ,34 and protect mice from IBD mediated by CD4+ T cells, natural killer (NK) cells and innate lymphoid cells (ILC).35–37 Interestingly, recent work also shows that neutrophils are important source of IL-22.14 As we found that CD177+ neutrophils expressed higher transcriptional levels of IL-22 than CD177− neutrophils from patients with CD and UC by RNA sequencing and qRT-PCR (figure 4A,F), we then investigated whether there was difference in IL-22 expression between mouse CD177+ and CD177− neutrophils. Neutrophils were isolated from the bone marrow of WT and CD177−/− mice and stimulated with IL-23 to test IL-22 production. We found that the production of IL-22 was markedly increased at both mRNA and protein levels in WT neutrophils compared with that in CD177−/− neutrophils (figure 6A–C, see online supplementary figure S9A). However, there was no significant difference of IL-22 secretion in CD4+ T cells and ILCs from colonic mucosa of WT and CD177−/− mice (see online supplementary figure S9B–E). Interestingly, we found that expression of zonula occludens (ZO)-1 was markedly decreased in IECs from CD177−/− mice compared with that in WT mice after DSS exposure (figure 6D), while expression of Reg3β and Reg3γ, an antimicrobial proteins produced by IECs, which can kill bacteria,38 was significantly increased in IECs from WT mice than those from CD177−/− mice (figure 6E,F), which was consistent with IL-22 production in neutrophils. We next asked whether CD177+ neutrophils could maintain intestinal barrier integrity. WT and CD177−/− mice were fed with fluorescein isothiocyanate (FITC)-dextran (500 mg/kg) 4 hours before anesthetisation on day 10 as described previously,39 ,40 and the levels of FITC-dextran in sera were detected by ELISA. As shown in figure 6G, CD177−/− mice displayed significantly higher serum levels of FITC-dextran compared with WT mice after DSS exposure. Next, we detected serum level of LPS to investigate microbial dissemination in sera, and found that its level was highly increased in CD177−/− mice after DSS exposure (figure 6H), indicating an increased bacterial load in CD177−/− mice and an important role of CD77 in maintaining intestinal barrier function on DSS treatment.

CD177+ neutrophils produce high levels of interleukin (IL)-22 and maintain epithelial barrier function. Neutrophils (2×106) isolated from the bone marrow of wild-type (WT, n=12) and CD177−/− (n=12) mice with or without dextran sulfate sodium (DSS) insult were first stimulated with lipopolysaccharide (LPS) (200 ng/mL) for 2 hours, and then cultured under stimulation with IL-23 (10 ng/mL) for further 4 hours. (A) Expression of IL-22 mRNA was examined by quantitative RT-PCR (qRT-PCR) and normalised to GAPDH. (B) After the stimulation of LPS and IL-23, neutrophils (2×106) were incubated in culture medium alone at 37°C in a humidified 5% CO2 incubator for 24 hours. Supernatants were collected for the detection of IL-22 using ELISA. (C) After the stimulation of LPS and IL-23, neutrophils (2×106) stimulated with PMA (50 ng/mL) and ionomycin (750 ng/mL) for 5 hours. Brefeldin A (3 μg/mL) was added into the culture medium for the last 3 hours. Expression of IL-22+ neutrophils in the bone marrow was determined by flow cytometry. (D−F) Intestinal epithelial cells (IECs) were isolated from WT and CD177−/− mice, and expression of ZO-1 (D), Reg3β (E) and Reg3γ (F) were examined by qRT-PCR. Gene expression was normalised to GAPDH. (G) WT (n=12) and CD177−/− (n=12) mice were induced colitis by 2% DSS. On day 10, all mice were administered with fluorescein isothiocyanate (FITC)-dextran (500 mg/kg) by oral gavage 4 hours before sacrifice. The sera were then obtained to detect level of FITC-dextran by fluorescence microplate at the excitation maximum of 490 nm and emission maximum of 520 nm. (H) Serum level of LPS was detected by spectrophotometry. *p<0.05, **p<0.01, ***p<0.001. Representative results are shown from three independent experiments.
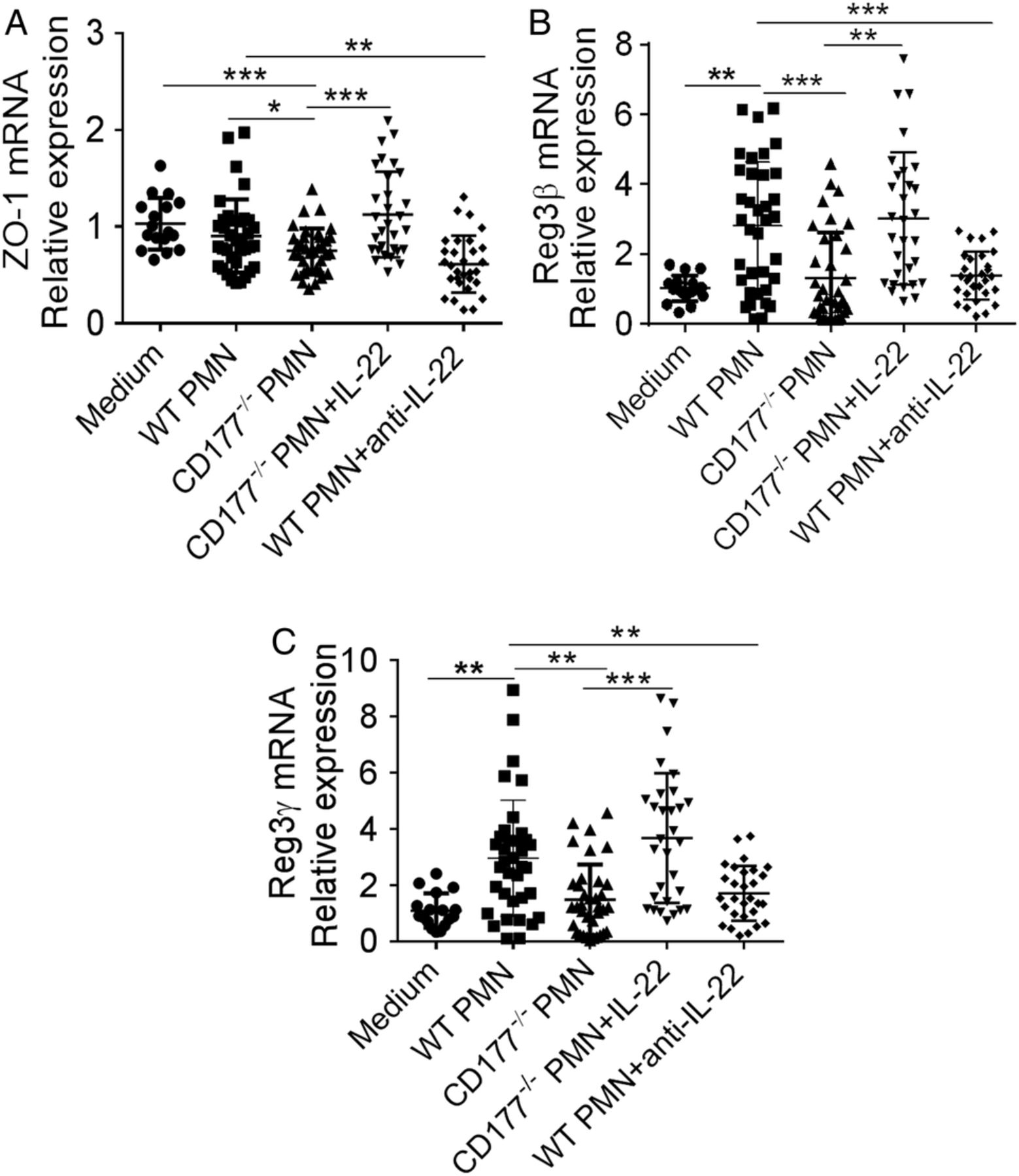
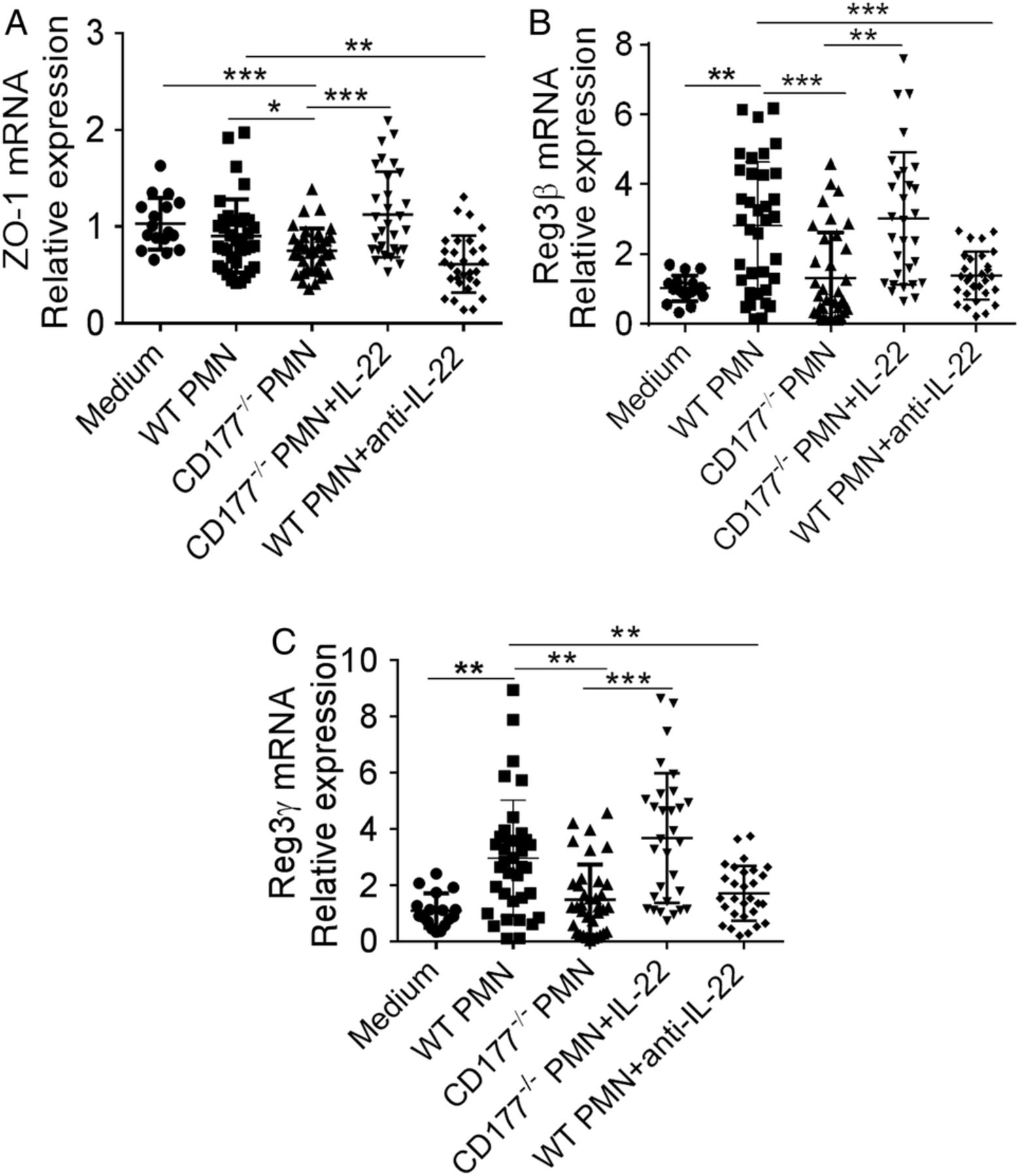
To further determine whether CD177+ neutrophils regulate intestinal epithelial cell barrier integrity through production of IL-22, we used a colon-derived epithelial cell line, MC38. Neutrophils were isolated from the bone marrow of WT and CD177−/− mice, stimulated with LPS for 2 hours and then cocultured with MC38 cells at the effector-to-target cell ratio of 5:1 for 24 hours. We found that level of ZO-1 was higher in MC38 cells cocultured with WT neutrophils than that in MC38 cells cocultured with CD177−/− neutrophils (figure 7A). The same pattern expression of Reg3β and Reg3γ was also observed (figure 7B,C). We then investigated whether increased expression of ZO-1 in MC38 cells cocultured with WT neutrophils was attributed to an increase of IL-22 production from WT neutrophils. As shown in figure 7, addition of exogenous IL-22 in MC38 cells cocultured with CD177−/− neutrophils significantly reversed expression of ZO-1, Reg3β and Reg3γ in MC38 cells. On the contrary, a neutralising antibody against IL-22 in MC38 cells cocultured with WT neutrophils resulted in markedly downregulated expression of ZO-1, Reg3β and Reg3γ in MC38 cells (figure 7). These data indicate that CD177+ neutrophils maintain epithelial barrier function through production of IL-22 and upregulate the levels of antimicrobial peptide produced by colonic epithelial cells.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
CD177+ neutrophils promote intestinal integrity. Neutrophils were isolated from the bone marrow of wild-type (WT) and CD177−/− mice after dextran sulfate sodium (DSS) insult, stimulated with lipopolysaccharide (LPS) (200 ng/mL) for 2 hours, and then cocultured with colon-derived MC38 epithelial cell line (1×105) at effector-to-target cell ratio of 5:1 in the presence of interleukin (IL)-23 (10 ng/mL) with or without IL-22 (10 ng/mL) or anti-IL-22 monoclonal antibody (1 μg/mL) for 24 hours. Expression of ZO-1 (A), Reg3β (B) and Reg3γ (C) in MC38 cells was examined by quantitative RT-PCR. Gene expression was normalised to GAPDH in each group. *p<0.05, **p<0.01, ***p<0.001. Data are pooled from three independent experiments.
Discussion
Neutrophils, a component of the innate immune system, have been shown to play a vital role in the maintenance of intestinal homeostasis.13 CD177 is a neutrophil-specific glycoprotein41 and has been shown to be involved in the pathogenesis of systemic vasculitis, SLE, asthma and polycythaemia vera.20 ,42 ,43 In this study, we demonstrated that CD177+ neutrophils are the functionally activated subset of neutrophils, which negatively regulate the pathogenesis of IBD.
During intestinal mucosal inflammation, neutrophils are critical component to maintain intestinal homeostasis. However, the roles of infiltrating neutrophils in the pathogenesis of IBD are still controversial as both pathological and beneficial roles being reported.13 It has been shown that neutrophils play a protective role in limiting intestinal mucosal inflammation. For example, depletion of neutrophils exacerbates 2,4,6-trinitrobenzene sulphonic acids (TNBS) or CD4+CD45RBhigh T cell-induced intestinal inflammation in mice.14 ,44 ,45 In our current study, we found that depletion of CD177, which is specifically expressed in neutrophils, aggravated DSS-induced colitis in mice, indicating a protective role of CD177+ neutrophils in IBD. The great importance of neutrophils in clearing invading microbes can be illustrated by depletion of neutrophils that leads to the enhanced translocation of bacteria in colitic mice.44 Neutrophils have been reported to have elegant defence mechanisms to eliminate extracellular microbes.13 When contacting with invading microbes, neutrophils phagocytose them.12 ROS are then generated in an oxidative burst by a NADPH oxidase, and antimicrobial proteins are released, including calprotectin, defensin and lactoferrin.28 Calprotectin, comprising S100A8 and S100A9, exhibits antimicrobial effects on bacteria, fungi and protozoa,46 which is reported to mediate NADPH oxidase activation and ROS production.47 ,48 Analysis of differential gene expression profiles between CD177+ and CD177− neutrophils has shown that functional differences were mainly reflected in defence response, inflammatory response, response to ROS and hydrogen peroxide in patients with CD or UC. Moreover, CD177+ neutrophils released high levels of ROS and antimicrobial peptides compared with CD177− neutrophils under inflammatory conditions. Therefore, our data illustrate that CD177+ neutrophils function as a protective subset in defence response, response to ROS and hydrogen peroxide during intestinal inflammation.
Evidences have demonstrated that highly activated neutrophils can eliminate microorganisms by releasing NET, which can imprison pathogens, prevent them from spreading, and facilitate the further phagocytosis of trapped microorganisms.12 NET is composed of nuclear component (DNA and histones) and proteins from granules (eg, MPO, elastase and lactoferrin),31 and has diverse functions, including antimicrobial activities, microbial recognition and tissue remodelling.11 Previous studies have shown that the efficiency of NET in bacterial trapping in blood was fourfold more than that of Kupffer cells during sepsis induced by Escherichia coli, and that blockade of NET formation resulted in E. coli dissemination to distant organs, indicating a critical role of NET in acute inflammation.12 Here, we found that NET formation was increased in CD177+ neutrophils compared with that in CD177− neutrophils, and that CD177+ neutrophils from patients with active IBD demonstrated more NET formation compared with controls. Thus, these data indicate that CD177+ neutrophils as functionally activated neutrophils function to eliminate invading microorganisms during acute inflammation in patients with CD or UC.
The hallmark of IBD is abnormal production of inflammatory cytokines, such as TNF-α, IFN-γ, IL-17A and TGF-β.49 Under spontaneous or stimulatory conditions, neutrophils are capable of producing several inflammatory cytokines and chemokines, such as IL-1β, IL-17A, IL-6, IFN-γ, TGF-β, CXCL1 and CCL4,11 which are further involved in the regulation of innate and adaptive immune responses to some extent. Neutrophil-derived cytokines have been studied to be highly associated with intestinal mucosal inflammation.11 ,13 ,50 In our study, we found that CD177+ neutrophils produced lower levels of IFN-γ, IL-17A and IL-6, but higher levels of IL-22 under stimulation with LPS. IL-22 has been reported to be highly upregulated in inflamed intestinal mucosa and sera from patients with IBD.34 IL-22R is specifically expressed in epithelial cells51 and IL-22 acts directly on epithelium, which is considered to be beneficial for intestinal epithelial barrier through enhanced mucus production, increased proliferation and migration of epithelial cells.33 ,34 Moreover, IL-22 contributes to the expression of Reg3β and Reg3γ antimicrobial proteins, which further promote bacterial elimination via direct microbicidal effects.38 During Citrobacter rodentium infection, IL-22 deficiency results in more weight loss and mortality, because of compromised intestinal integrity, decreased epithelial antimicrobial defence,38 suggesting a protective role of IL-22 against microbial infection. It has been reported that IL-22 is mainly produced by T helper 17, NK T, CD8 T and γδ T cells.35 ,52 ,53 However, neutrophils are found to be an important source of IL-22,14 ,15 and IL-22-producing neutrophils could enhance the production of epithelial microbial peptides, promote to restore epithelial integrity and contribute to resolution of colitis.15 In our current study, CD177+ neutrophils produced high levels of IL-22 and further contributed to increased production of Reg3β and Reg3γ antimicrobial proteins. However, without CD177+ neutrophils-derived IL-22, CD177−/− mice demonstrated an increased level of LPS in sera and compromised intestinal barrier. Collectively, these data highlight the important role of CD177+ neutrophils in protecting intestinal epithelial integrity through increased production of IL-22 during intestinal inflammation.
In summary, our studies reveal that CD177+ neutrophils, as functionally activated subset of neutrophils, play a protective role in the pathogenesis of IBD. Our study thus identify critical roles of CD177+ neutrophils in regulating intestinal mucosal inflammation through increased bactericidal activities, high level of IL-22 and less production of proinflammatory cytokines. Therefore, targeting CD177 or CD177+ neutrophils could provide a novel approach in the treatment of IBD.
References
Footnotes
*GZ and LY co-first authors.
Contributors ZL designed and conducted the experiments, acquired and analysed the data; GZ, LY and LF performed all experiments; WY, TY, YM, MC, KW and FC analysed the data; LF and WY contributed to the clinical data and specimens; GZ, YC and ZL wrote the manuscript. All authors discussed and revised the manuscript.
Funding This work was financially supported by grants from the National Natural Science Foundation of China (81630017, 81470822).
Competing interests None declared.
Patient consent Obtained.
Ethics approval Institutional Review Board for Clinical Research of the Shanghai Tenth People's Hospital of Tongji University.
Provenance and peer review Not commissioned; externally peer reviewed.