Article Text

Abstract
Objective In the tumour microenvironment, critical drivers of immune escape include the oncogenic activity of the tumour cell-intrinsic osteopontin (OPN), the expression of programmed death ligand 1 (PD-L1) and the expansion of tumour-associated macrophages (TAMs). We investigated the feasibility of targeting these pathways as a therapeutic option in hepatocellular carcinoma (HCC) mouse models.
Design We analysed the number of tumour-infiltrating immune cells and the inflammatory immune profiles in chemically induced liver tumour isolated from wild-type and OPNknockout (KO) mice. In vitro cell cocultures were further conducted to investigate the crosstalk between TAMs and HCC cells mediated by OPN, colony stimulating factor-1 (CSF1) and CSF1 receptor (CSF1R). The in vivo efficacy of anti-PD-L1 and CSF1/CSF1R inhibition was evaluated in OPN overexpressing subcutaneous or orthotopic mouse model of HCC.
Results The numbers of TAMs, as well as the expression levels of M2 macrophage markers and PD-L1 were significantly decreased, but the levels of cytokines produced by T-helper 1 (Th1) cells were upregulated in tumour tissues from OPN KO mice compared with that from the controls. In addition, we observed a positive association between the OPN and PD-L1 expression, and OPN expression and TAM infiltration in tumour tissues from patients with HCC. We further demonstrated that OPN facilitates chemotactic migration, and alternative activation of macrophages, and promotes the PD-L1 expression in HCC via activation of the CSF1-CSF1R pathway in macrophages. Combining anti-PD-L1 and CSF1R inhibition elicited potent antitumour activity and prolonged survival of OPNhigh tumour-bearing mice. Histological, flow cytometric and ELISA revealed increased CD8+ T cell infiltration, reduced TAMs and enhanced Th1/Th2 cytokine balance in multiple mouse models of HCC.
Conclusions OPN/CSF1/CSF1R axis plays a critical role in the immunosuppressive nature of the HCC microenvironment. Blocking CSF1/CSF1R prevents TAM trafficking and thereby enhances the efficacy of immune checkpoint inhibitors for the treatment of HCC.
- hepatocellular carcinoma
- tumor microenvironment
- immune checkpoint blockade
- anti-PD-L1
Statistics from Altmetric.com
Significance of this study
What is already known on this subject?
Hepatocellular carcinoma (HCC) remains one of the most lethal human malignancies with limited effective treatment options. Targeting tumour-associated macrophages (TAMs) is a promising strategy.
Checkpoint blockade immunotherapy has limited efficacy in patients with HCC.
Critical drivers of immune escape in tumour microenvironment include the oncogenic activity of the HCC cell-intrinsic osteopontin (OPN), the expression of programmed death ligand 1 (PD-L1), and the expansion of TAMs.
What are the new findings?
Chemically induced tumours from OPN knockout mice had reduced numbers of TAMs, reduced expression of markers of M2 macrophage and PD-L1 and increased levels of cytokines produced by T-helper 1 (Th1) cells compared with tumours from control mice.
OPNhigh HCC facilitates M2-like polarisation of macrophages and promotes PD-L1 expression via the activation of the colony stimulating factor-1 (CSF1)/CSF1 receptor (CSF1R) pathway in macrophages, which enhances the production of immunosuppressive cytokines and drives HCC metastasis through successful evasion of Th1-dependent tumour elimination.
Inhibition of the CSF1/CSF1R pathway reduces macrophage recruitment and M2 phenotype polarisation, which subsequently activates CD8+ T cells and sensitises immune-dependent OPNhigh HCC to anti-PD-L1 immune checkpoint blockade.
How might it impact on clinical practice in the foreseeable future?
Strategies to block CSF1 signalling via CSF1R on TAMs might increase the efficacy of immune checkpoint inhibitors in patients with OPNhigh HCC.
Introduction
Hepatocellular carcinoma (HCC) remains one of the most lethal human malignancies with limited treatment options.1 Moreover, only a small number of patients with HCC are eligible for curative treatments, such as surgical resection, liver transplantation and ablation.2 Many molecular targeted therapies, such as sorafenib, lenvatinib, regorafenib and cabozantinib, have been approved for the treatment of advanced HCC.3–6 Moreover, the US Food and Drug Administration (FDA) approved antiprogrammed death receptor 1 (anti-PD-1) antibody nivolumab and pembrolizumab for treatment of HCC who have been previously treated with sorafenib.7 8 However, due to frequent drug resistance, only modest improvement in survival has been observed with these therapies. Thus, identifying novel therapies to improve long-term patient outcome are urgently needed.
Recent advances in immunotherapy has transformed cancer treatment by blocking immune checkpoints, for example, programmed death ligand 1 (PD-L1), programmed death receptor 1 (PD-1) or cytotoxic T-lymphocyte associated protein 4 (CTLA-4), to unleash antitumour immune response.9 PD-L1 is one of two ligands for PD-1 and expressed on the surface of cancer cells, dendritic cells and macrophages.10–12 Ligation of PD-L1 on cancer cells to PD-1 expressed on T cells induces T cell exhaustion and blocks further immune rejection, bypassing immune surveillance. Upregulation of intratumoural PD-L1 has been observed in patients with HCC and reported to correlate with poor prognosis.13
Tumours that arise in immunosufficient hosts are often poorly immunogenic as a result of immunoediting, a dynamic process of immunosurveillance to immune escape.14 Although multiple factors have been shown to contribute to the loss of immunogenicity, one dominant driver is the recruitment of suppressive immune cells or the production of immunosuppressive factors.15 Among the immunosuppressive cells, tumour-associated macrophages (TAMs) are believed to play critical roles in immune suppression and contribute to HCC malignant progression.16–18 Accumulated evidence suggests that increased density of TAMs correlates with early recurrence, leading to poorer survival in patients with HCC.19 20 More recently, PD-L1/PD-1 ligation between tumour and TAMs is also reported to inhibit of macrophage-mediated phagocytosis of tumour cells.21 In this regard, therapeutics that can reprogramme TAMs by targeting key regulators for M2 phenotype polarisation, which is associated with immunosuppression and tumour progression, might overcome immunosuppression and enhance response to anti-PD-1/PD-L1 therapy.
Previous studies by our group and others demonstrated that osteopontin (OPN), a prominent tumour-sustaining inflammatory mediator, is associated with tumour progression, metastasis and immunosuppression.22–25 However, the relationship between OPN and the immunosuppressive phenotype in HCC microenvironment remains unclear. Here, we report that OPN facilitates chemotactic migration, and M2-like polarisation of macrophages, and promotes the expression of PD-L1 in HCC via activation of the colony stimulating factor-1 (CSF1)–CSF1 receptor (CSF1R) pathway in macrophages, which leads to increased production of immunosuppressive cytokines. Moreover, activation of the CSF1/CSF1R pathway also blocked Th1-mediated immune response and other classical anti-tumour immune effector pathways. Notably, our findings indicated that blocking the CSF1/CSF1R pathway prevented TAM trafficking and enhanced the efficacy of immune checkpoint-based therapies in HCC by OPN overexpression and suggested that CSF1R inhibitor may be used as an adjuvant for immune checkpoint-based treatment of HCC.
Methods
Cell cultures
HEK 293 T cells, human HCC cells (Hep3B and HepG2), mouse HCC cell lines (Hepa1-6 and H22) and human monocytic THP-1 cells were purchased from the Institute of Biochemistry and Cell Biology, Chinese Academy of Science (Shanghai China). Human HCC cell lines, MHCC97H and HCC-LM3, were obtained from the Liver Cancer Institute at Fudan University (Shanghai, China) as previously described.26
Animal studies
Male OPN knockout (KO) mice (C57BL/6 background) and their corresponding wild-type (WT) littermates were kindly provided by Professor Jin-Hong Chen at Fudan University (Shanghai, China) and have been previously described.27 Fourteen-day-old OPN KO and WT mice administered 25 mg/kg diethylnitrosamine (DEN; Sigma Aldrich, St. Louis, Missouri, USA) via intraperitoneal (i.p.) injection to induce hepatocarcinogenesis. Two weeks after DEN treatment, mice were administered 0.5 µL/g carbon tetrachloride (CCl4) dissolved in corn oil via i.p. injection once per week for 20 weeks as previously described.28
For additional methods can be found in the online supplementary methods.
Supplemental material
Results
OPN deletion shifts the immune and inflammatory profiles at the tumour site
To determine the effects of OPN on HCC immune microenvironment in vivo, we first established a classical DEN/CCl4-induced mouse of HCC with OPN KO using a previously established protocol for chemically induced carcinogenesis.28 Compared with the WT counterparts, OPN KO mice had less HCC incidence, smaller histological lesions, enhanced apoptosis, reduced tumour vascularisation and less lung metastases (figure 1A and online supplementary figure 1A–G).
Supplemental material

OPN deletion suppresses infiltration of TAMs and HCC PD-L1 expression and shifts the immune and inflammatory profile in the liver of mice undergoing DEN/CCl4-induced hepatocarcinogenesis. (A) Top: schematic representation of DEN/CCl4-induced HCC model. Fourteen-day-old mice were administered DEN (25 mg/kg) by i.p. injection. After 2 weeks, mice were then administered CCl4 (0.5 mL/kg, dissolved in corn oil) by i.p. injection. Bottom: representative gross appearance of livers from 12-month-old DEN/CCl4-treated male WT (n=6) and OPN KO (n=6) mice. (B) Left: 12-month-old DEN/CCl4-treated mice. Tumour-infiltrating immune cells and inflammatory cells, including T cells (CD4+ and CD8+), T regulatory (Treg) cells (CD4+CD25+Foxp3+), natural killer (NK) cells (NK1.1+), B lymphocytes (CD19+) and TAMs (F4/80+CD11b+) were detected by flow cytometry in liver tumour tissues. Right: representative flow cytometric analysis showing the proportion of CD4+, CD8+ T cells and TAMs in liver tumour tissues of mice. (C) Heatmap of a selected list of genes showing the fold change of expression in OPN KO mice normalised to those from WT mice. Each data was replicated triply. (D) Relative expression of the indicated genes by qPCR. Data depict the mean±SEM (n=6) and are representative of three independent experiments. (E) Representative images of PD-L1, F4/80 and CD206 immunostaining in liver tissue sections of WT and OPN KO. Scale bar: 100 µm. (F) Flow cytometric analysis of TAMs (F4/80+CD11b+) and neoplastic PD-L1 expression (CD45−PD-L1+) in liver tumour tissues from WT and KO mice at the indicated time points after DEN injection. *P<0.05, **p<0.01 and ***p<0.001 versus WT, Student’s t-test. CCl4, carbon tetrachloride; HCC, hepatocellular carcinoma; KO, knockout; OPN, osteopontin; PD-L1, programmed death ligand 1; TAMs, tumour-associated macrophages; WT, wild-type.
The percentage of tumour-infiltrating immune and inflammatory cells29 and the expression of immune checkpoints and mediators of inflammation and immunity in DEN/CCl4-induced mouse liver tumour tissues were assessed by flow cytometric analysis. We observed a significant reduction of TAMs in OPN KO tumours compared with WT tumours (figure 1B and online supplementary figure 2A). The percentage of other immune cells, for example, CD8+ T cells, CD4+ T cells, regulatory T (Treg) cells, B cells and natural killer cells, did not change significantly (figure 1B). Similarly, OPN deletion also resulted in a significant decrease in the percentage of blood monocyte but did not induce a significant increase in the numbers of CD8+ T and CD4+ T cells in DEN/CCl4-treated mice (see online supplementary figure 2B). To explore the possibility of T cell checkpoint inhibitors for the treatment of HCC, we evaluated expression of the PD-1, PD-L1, CTLA-4, T cell immunoglobulin and mucin-domain containing-3 (TIM-3) and lymphocyte activation gene 3 in OPN KO tumours and WT tumours. The expression of PD-L1 mRNA was significantly decreased on OPN deletion, whereas no appreciable changes in the expression level of other immune checkpoints were observed (figure 1C,D). In contrast, the mRNA levels of several known antitumour immune mediators indicative of antitumour Th1 immunity, such as interferon γ (IFN-γ), tumour necrosis factor α (TNFα), C-X-C motif chemokine ligand 10 (CXCL10) and interleukin (IL)-12b,30 were markedly increased in OPN KO tumours (figure 1C,D). Likewise, mRNA levels of type I IFN-stimulated genes, IFIT1 and IFIT2, were also elevated in OPN KO tumours, indicating enhanced type I IFN (IFN-α/β) signalling.31 We also observed a reduction in the expression of Th2 cytokines, for example, IL-6, CSF1, and CXCL1, as well as markers associated with M2 macrophage polarisation, for example, type I arginase (Arg1), in OPN KO tumours (figure 1C,D and online supplementary figure 2C). Immunohistochemistry (IHC) staining data indicated a decrease in TAMs (F4/80) and expression of neoplastic PD-L1 in OPN KO tumours (figure 1E and online supplementary figure 2D). To further validate the role of OPN in regulating HCC immune microenvironment, during hepatocarcinogenesis in vivo, we analysed variations in TAM infiltration and neoplastic PD-L1 expression at different time points of DEN/CCl4-induced liver tumourigenesis in both WT and OPN KO mice. Flow cytometric analysis revealed a progressive increase in the fraction of infiltrating TAMs (F4/80+CD11b+) and neoplastic PD-L1 expression (CD45−PD-L1+) in the liver of WT mice treated with DEN/CCl4 at months 6, 9 and 12 (figure 1F). Moreover, similar changes in hepatic PD-L1 expression were observed at months 9 and 12 in DEN/CCl4-induced model of HCC (see online supplementary figure 2E,F). However, the PD-L1 expression on the infiltrating haematopoietic cells (CD45+/PD-L1+) were not significantly changed on OPN deletion (online supplementary figure 2G). These findings suggest OPN deletion attenuates PD-L1 expression, and TAM infiltration, and enhances the anti-tumour immune signalling in HCC.
To investigate the effects of HCC-derived OPN on the inflammatory response in vivo, we orthotopically inoculated C57BL/6 mice with control or Hepa1-6-shOPN (OPN knockdown by shRNA) cells. Knockdown of OPN in Hepa1-6 cells suppressed tumour growth and lung metastasis and increased the overall survival compared with the control (see online supplementary figure 3A–G). In addition, we also measured TAM infiltration and neoplastic PD-L1 within tumours by flow cytometry. Similar to that observed in the DEN/CCl4-induced mice, knockdown of OPN in HCC cells significantly decreased the population of TAMs (F4/80+CD11b+) and CD45−PD-L1+ cells (figure 2A and online supplementary figure 4A). However, there was no significant alteration in the percentage of PD-1 on CD8+ T cells (see online supplementary figure 4A). The decreased density of F4/80, CD206 and PD-L1 was consistently observed in OPN knockdown tumours by IHC staining (figure 2B and online supplementary figure 4B). In particular, the decreased expression of M2 markers, Arg1, CD206 and IL-10 was accompanied by enhanced expression of M1 markers, iNOS, IL-1b and IL-12b, in HCC cells with OPN knockdown (figure 2C). Moreover, consistent with the above results (figure 1D), we also observed decreased expression of PD-L1 and elevated expression of genes that are associated with effector T cell differentiation (Tbx21)32 and antitumour Th1 immunity, such as IFN-γ, TNFα and CXCL10 (see online supplementary figure 4C).30 Importantly, compared with control tumours, OPN knockdown tumours exhibited an apparent shift in the immune and inflammatory profiles towards an M1-like activation and increased expression of antitumour immune mediators. Together, these results suggest that Hepa1-6 HCC-derived OPN drives the expression of immune checkpoint PD-L1 and multiple tumour-promoting cytokines in the local tumour microenvironment, while blocking the activation of Th1 immunity and other antitumour immune effector pathways.

The effects of OPN on TAM infiltration and PD-L1 expression in HCC. C57BL/6 mice were inoculated with Hepa1-6-shcontrol or Hepa1-6-shOPN cells, and the expression of neoplastic PD-L1 and TAM infiltration was determined. (A) Flow cytometric analysis of the infiltrated TAMs (F4/80+CD11b+) and neoplastic PD-L1 expression (CD45−PD-L1+) in orthotopic tumours from the indicated groups. (B) TAM infiltration and neoplastic PD-L1 expression in orthotopic tumours of the indicated groups as determined by F4/80, CD206 and PD-L1 immunostaining. Representative IHC images of the indicated groups are shown. Scale bar: 100 µm. (C) Expression of markers associated with M1 and M2 polarisation in tumours of the indicated groups. (D) Flow cytometric analysis of infiltrated TAMs and neoplastic PD-L1 expression isolated from human HCC tumour tissues (n=20). Top: the percentage of TAMs (CD68+CD163+) and statistical analysis of TAMs in HCC with high or low OPN expression. Bottom: the percentage of neoplastic PD-L1 (CD45–PD-L1+) and statistical analysis of neoplastic PD-L1 expression in HCC with high or low OPN expression. (E) Representative images of IHC staining of OPN, PD-L1 and TAM infiltration in HCC tissues. Scale bar: 100 µm. *P<0.05, **p<0.01 and ***p<0.001, Student’s t-test. HCC, hepatocellular carcinoma; OPN, osteopontin; PD-L1, programmed death ligand 1; TAMs, tumour-associated macrophages.
Expression of OPN associates with neoplastic PD-L1 expression and TAM infiltration and predicts poor prognosis in patients with HCC
The results shown above prompted us to investigate the association between OPN, neoplastic PD-L1 and TAM infiltration in HCC patient tissues, then we performed flow cytometric analysis of fresh human HCC tissues and compared the levels of intratumoural OPN, neoplastic PD-L1 and infiltrated TAMs. The results showed that neoplastic PD-L1 expression (CD45−PD-L1+) and the levels of TAMs (CD68+CD163+) were significantly higher in OPNhigh than in OPNlow HCC tissues (figure 2D).
Consistent with the above results, IHC staining revealed a positive association between intratumuoral OPN expression and neoplastic PD-L1 expression or TAM infiltration (figure 2E and see online supplementary table1). In addition, both OPN expression and TAM infiltration were significantly associated with the presence of tumour thrombus, poorer tumour differentiation and higher TNM stage (see online supplementary tables 2–4). HCC patients with high OPN expression and PD-L1 had shorter overall survival and higher recurrence rates than patients with low OPN and PD-L1 expression (see online supplementary figure 4D,E). Moreover, the highest recurrence rates and the lowest overall survivals were observed in patients with both high OPN and high PD-L1 expression (see online supplementary figure 4F). Similarly, intratumoural TAM infiltration was associated with poorer prognosis (see online supplementary figure 4D,E), and the poorest prognosis was observed among patients with both high OPN and high TAM infiltration (see online supplementary figure 4G). Multivariate analysis indicated that overexpression of PD-L1 was an independent risk factor for recurrence and survival (see online supplementary tables 5 and 6). Finally, we interrogated publicly available gene expression data in human HCC found that OPN expression had a weak (r<0.4) but significant (p<0.05) correlation with TAM infiltration (see online supplementary figure 4H,I). These findings suggest OPN-mediated expression of PD-L1 and TAM infiltration has the potential to serve as a prognostic indicator for patients with HCC.
Supplemental material
OPN induces chemotactic migration and M2-like polarisation of macrophages and promotes the PD-L1 expression in HCC
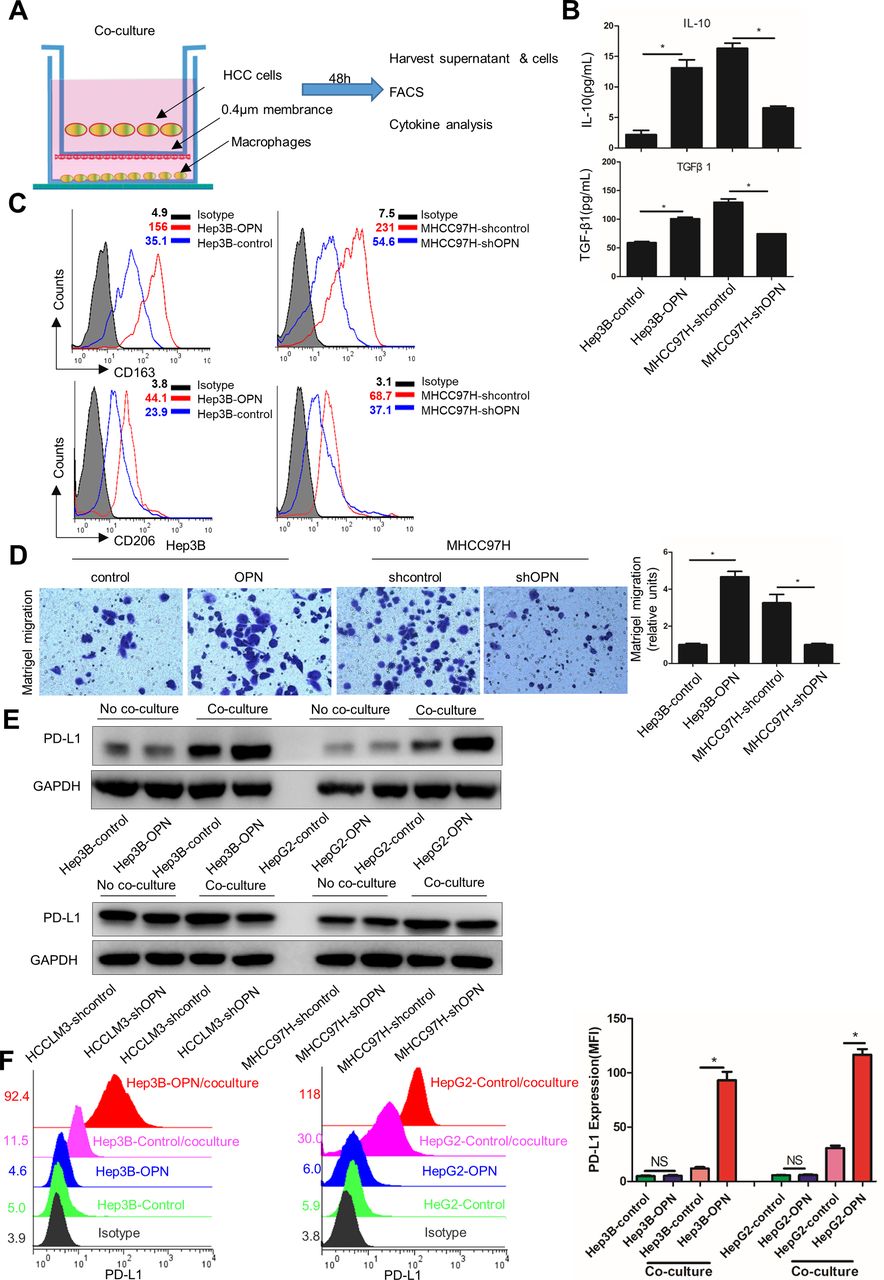
To further validate the association between OPN expression and inflammatory response, we first established an in vitro coculture system by culturing HCC cells with different OPN expression levels with phorbol-12-myristate-13-acetate (PMA)-treated THP-1 cells, which induces monocytic THP-1 cell line into monocyte-derived macrophages. We then assessed the M2-like polarisation of macrophages and expression of PD-L1 in these cells (figure 3A). Macrophages cocultured with OPNhigh HCC cells (Hep3B-OPN and MHCC97H-shcontrol) produced significantly more tumour-promoting cytokines, such as IL-10 and transforming growth factor-β1 (TGF-β1)33 and exhibited a CD163high/CD206high phenotype,34 which is similar to TAMs phenotype, compared with those cocultured with OPNlow HCC cells (Hep3B-control and MHCC97H-shOPN) (figure 3B,C and online supplementary figure 5A). Consistent with the changes in cytokine secretion and cell surface markers, OPNhigh-HCC-stimulated macrophages also exhibited enhanced expression of M2 markers, IL-4, CCL22 and CD206, with decreased expression of M1 markers, IL-1b, IL-12b and TNFα, indicating functional activation of TAMs (see online supplementary figure 5B). To explore whether OPN affects chemotactic migration of macrophages, we conducted a migration assay using PMA-treated THP-1 cells. As shown in figure 3D, the supernatants from OPN-overexpressing HCC cells cocultured with macrophages induced the chemotactic migration of THP-1 cells treated with PMA. These results suggest that OPN induces the migration and M2-like polarisation of macrophages in vitro.

OPN facilitates chemotactic migration, M2-like polarisation of macrophages and upregulation of PD-L1 in HCC. (A) Schematic showing the HCC cells cocultured with human monocytic-leukemia cells (THP-1) stimulated by PMA in a transwell apparatus of 0.4 µm pore size. After 48 hours, HCC cells and macrophages were collected for subsequent analyses described in figure parts B–D. (B) Supernatants were analysed by ELISA for the levels of IL-10 and TGF-β1. (C) Flow cytometric analysis of CD163 and CD206 expression on macrophages. (D) PMA-stimulated THP-1 cells were subjected to migration assays towards the supernatants of HCC cells cocultured with macrophages. (E and F) Macrophages were cocultured with Hep3B-OPN or HepG2-OPN (Hep3B-control or HepG2-control used as control) for 48 hours. Western blot analysis (E) and flow cytometric analysis (F) of PD-L1 expression in HCC cells with different OPN expression and with or without cocultured macrophages. *P<0.05, Student’s t-test. Data represent the means±SEM of triplicate experiments. HCC, hepatocellular carcinoma; IL, interleukin; OPN, osteopontin; PD-L1, programmed death ligand 1; PMA, phorbol-12-myristate-13-acetate; TGF-β1, transforming growth factor-β1.
We initially speculated that the mechanism underlying the association between the OPN expression and PD-L1 might involve a direct regulation of PD-L1 expression by OPN. However, we did not observe significant changes in PD-L1 protein expression in OPN-overexpressing or silenced HCC cells (see online supplementary figure 5C). These results strongly suggested that OPN may influence PD-L1 expression via macrophages in HCC microenvironment. To test this hypothesis, we evaluated the potential effect of macrophages on the expression of PD-L1 in HCC cells by western blot and flow cytometric analyses. We found that PD-L1 expression was significantly higher in OPNhigh HCC cells (Hep3B-OPN and HepG2-OPN) than in OPNlow cells (Hep3B-control and HepG2-control) after coculture with macrophages (figure 3E,F and online supplementary figure 5D). Collectively, these data suggest that OPN from HCC induces M2-like macrophage phenotype, which in turn promotes the PD-L1 expression in HCC.
Blockade of the CSF1-CSF1R axis reverses OPN-induced chemotactic migration and M2-like polarisation of macrophages and upregulation of PD-L1 in HCC
To identify the cytokines secreted by macrophages that is activated by OPNhigh HCC cells, we compared the cytokine profiles of the supernatants from Hep3B-control/macrophage and Hep3B-OPN/macrophage cocultures using a RayBio Human Cytokine Antibody Array. The levels of six cytokines, epidermal growth factor, macrophage colony-stimulating factor (CSF1), granulocyte-macrophage colony-stimulating factor (CSF2), tumour necrosis factor β, stem cell factor and monocyte chemotactic protein-3, were significantly higher in the supernatant of Hep3B-OPN/macrophage coculture compared with that of Hep3B-control/macrophage (see online supplementary figure 5E and table 7). CSF1, a ligand for CSF1 receptor (CSF1R), plays a pivotal role in monocyte/macrophage generation, survival and function.35 Previously, we reported a critical role of systemic CSF1 expression in HCC patients with metastasis.36 Because CSF1 expression was markedly decreased in OPN-KO tumours (figure 1C,D), we further evaluated the CSF1 levels in HCC/macrophage coculture evaluated by ELISA. We found that OPNhigh HCC/macrophage coculture exhibited about a 1.84–7.06-fold increase in the levels of CSF1, compared with control whereas the OPNlow HCC macrophage coculture exhibited decreased CSF1 levels (see online supplementary figure 5F).
To determine whether OPN-activated macrophages require the CSF1/CSF1R pathway, we performed an in vitro coculture assay and chemotactic migration analysis. We found that M2-like polarisation and macrophage migration were enhanced when cocultured with MHCC97H-shOPN cells treated with recombinant CSF1 (rhCSF1) or by lentivirus (LV)-mediated CSF1 overexpression (figure 4A–C and online supplementary figure 6A–C). In contrast, inhibition of CSF1 by either CSF1 shRNA (LV-shCSF1) or CSF1R inhibitor significantly decreased M2-like polarisation and migratory abilities of macrophages induced by OPN overexpression in Hep3B cocultures. The addition of CSF1 neutralising antibody or CSF1R inhibitor abrogated the upregulation of PD-L1 expression in Hep3B-OPN and HepG2-OPN cells as shown in the in vitro coculture assays (figure 4D,E, and online supplementary figure 6D). Given the importance of IFN-γ signalling pathway in PD-L1 expression, we evaluate the effects of CSF1 and IFN-γ on PD-L1 expression. As shown in online supplementary figure 6E,F, both CSF1 and IFN-γ could induce a significant increase of PD-L1 expression in Hep3B. In addition, their combination has a significant synergistic effect on PD-L1 expression in Hep3B cells. These findings suggest that the CSF1/CSF1R pathway likely plays a critical role in OPN-induced chemotactic migration and M2-like polarisation of macrophages and upregulation of PD-L1 in HCC.

CSF1/CSF1R blockade reverses OPN-induced migration, M2-like polarisation of macrophages and upregulation of PD-L1 in HCC. (A–C) Macrophages were infected with lentivirus (LV)-shCSF1 and LV-CSF1 for 24 hours or treated with CSF1R inhibitor (10 µM) and recombinant human CSF1 (rhCSF1). Macrophages were then cocultured with Hep3B-OPN or MHCC97H-shOPN cells for 48 hours. (A) Flow cytometric analysis of CD163 and CD206 expression on macrophages. (B) IL-10 and TGF-β1 in the supernatants detected by ELISA. *P<0.05, Student’s t-test. Data represent the means±SEM of three independent experiments. (C) Supernatants were collected for analysis for the effects of CSF1/CSF1R pathway on the migration of PMA stimulated THP-1 cells. (D and E) Macrophages were preincubated with CSF1R inhibitor or CSF1 blocking antibody and then cocultured with Hep3B-OPN or HepG2-OPN (Hep3B-control or HepG2-control) cells for 48 hours. Western blot (D) and flow cytometric (E) analysis of PD-L1 expression in the indicated HCC cells. CSF1, colony stimulating factor-1; CSF1R, CSF1 receptor; LV-CSF1, lentivirus-mediated colony stimulating factor-1; HCC, hepatocellular carcinoma; IL, interleukin; OPN, osteopontin; PD-L1, programmed death ligand 1; TGF-β1, transforming growth factor-β1.
OPN induces macrophage-mediated secretion of CSF1 via the PI3K/AKT/NF-κB/p65 pathway
To examine whether OPN induces CSF1 secretion in macrophages, we treated macrophages with recombinant human OPN and found that OPN significantly increased CSF1 expression in a dose-dependent manner (figure 5A). Next, we asked whether OPN induces CSF1 expression through transcriptional regulation. We compared the CSF1 promoter activity in OPN-treated macrophages and control cells, which indicated significantly higher CSF1 promoter activity after OPN treatment (figure 5B). Bioinformatics analysis identified two putative NF-κB/p65-binding sites within the CSF1 promoter region (see online supplementary figure 7A,B), suggesting that the NF-κB/p65 binding sites may be required for the activation of the CSF1 promoter by OPN, since it is known that OPN can activate NF-κB/p65 pathway.23 To test this hypothesis, we established a luciferase reporter with the CSF1 promoter to evaluate its transactivation by OPN. Both promoter activity and ELISA analyses indicated that NF-κB/p65 knockdown significantly decreased OPN-induced CSF1 secretion (figure 5C). Likewise, NF-κB/p65 pathway inhibitor, IMD-0354, also significantly suppressed OPN-induced CSF1 promoter transactivation and CSF1 secretion (figure 5D). To further validate NF-κB/p65 activation in OPN-treated macrophages, we examined nuclear expression of phospho-p65 (p-p65) by western blot and immunofluorescent staining. Consistent with the enhanced NF-κB/p65 promoter activity, translocation of nuclear p-p65 was also induced in OPN-treated macrophages (see online supplementary figure 7C,D).

OPN increases macrophage secretion of CSF1 via the PI3K/AKT/NF-κB/p65 pathway. (A) Macrophages were treated with various concentrations of OPN for 48 hours. CSF1 expression was detected by qPCR (left) and ELISA (right). (B) Macrophages were transiently transfected with the CSF1 promoter luciferase reporter and were treated with or without OPN. Luciferase activity was detected 48 hours after OPN treatment. (C) Macrophages were transfected with NF-κB/p65 shRNA or control shRNA, and then treated with or without OPN for 48 hours. After OPN treatment, the promoter activity and secretion of CSF1 were measured by luciferase reporter assay and ELISA, respectively. (D) Macrophages were treated with or without OPN in the presence of NF-κB/p65 inhibitor IMD-0354. The promoter activity and secretion of CSF1 were measured by luciferase reporter assay and ELISA, respectively. (E) Macrophages were pretreated with inhibitors against PI3K (LY294002), ERK1/2 (U0126) or p38 (SB203580) followed by OPN treatment. Left: expression of phosphorylated and total AKT, ERK and p38 were analysed by western blot. Right: ELSA to determine CSF1 secretion from the indicated treatment groups. (F) Chromatin immunoprecipitation-qPCR of NF-κB/p65 on the CSF1 promoter in the presence OPN and PI3K inhibitor. *P<0.05, Student’s t-test. Data represent means±SEM of triplicate experiments. CSF1, colony stimulating factor-1; OPN, osteopontin.
To determine which signalling pathway is involved in OPN-induced CSF1 secretion in macrophages, we treated macrophages with inhibitors of the PI3K, ERK and p38 kinases, as OPN can induce the activation of these pathways in a dose-dependent manner (see online supplementary figure 7E). Pretreatment of macrophages with the PI3K inhibitor but not ERK or p38 inhibitor reduced OPN-induced CSF1 secretion and translocation of nuclear p-p65 (figure 5E and online supplementary figure 7F). Chromatin immunoprecipitation-qPCR assay showed significantly reduced binding of NF-κB/p65 to the CSF1 promoter only in the presence of PI3K inhibitor but not ERK or p38 inhibitor (figure 5F). Collectively, these results suggest that OPN induces CSF1 release from macrophages through the PI3K/AKT/NF-κB/p65 pathway.
Inhibition of CSF1R enhances the efficacy of anti-PD-L1 blockade in OPNhigh tumour-bearing mice
Given the ability of TAMs to limit antitumour T cell response, we sought to determine whether targeting TAMs by CSF1/CSF1R inhibitor enhances HCC response to anti-PD-L1 blockade; we established subcutaneous Hepa1-6-OPN HCC tumours with OPN overexpression in C57BL/6 mice. Once tumours were palpable, mice were treated with isotype control antibody, anti-PD-L1 and CSF-1R inhibitor PLX3397, which has been granted by FDA for tenosynovial giant cell tumour,37 or the combination of anti-PD-L1 and PLX3397 until study endpoint (online supplementary figure 8A). The combination of anti-PD-L1 and PLX3397 significant inhibited tumour growth compared with control or single agent alone (figure 6A–C). In addition, no significant changes in average mice body weight or toxicity in the liver and kidney were observed (online supplementary figure 8B,C). To further validate the efficacy of the combination treatment, we established an orthotopic OPN-overexpressing H22 HCC mouse model. At 2 weeks after implantation, mice were administered anti-PD-L1, PLX3397 or the combination until the study endpoint (see online supplementary figure 8D). We observed significant tumour regression, reduced number of lung metastases and prolonged survival for the combination compared with that of control or single agent alone (figure 6D–H). Then, we performed the same combinatory treatment in the subcutaneous Hepa1-6-control HCC tumours and found that the PLX3397 or anti-PD-L1 alone induced significantly higher antitumour activity than control (p<0.05). Moreover, PLX3397 enhanced an efficacy of anti-PD-L1 treatment as well but less effective than Hepa1-6-OPN tumours (online supplementary figure 8E). These findings demonstrate the potential to improve treatment efficacy by blocking the CSF1/CSF1R and PD-1/PD-L1 axes in OPNhigh tumours.

Inhibition of OPN/CSF1/CSF1R signalling sensitises anti-PD-L1 blockade in HCC mouse models. (A–C) OPN-overexpressing Hepa1-6-OPN mouse HCC cells were subcutaneously injected into C57BL/6 mice. When tumours reached 50 mm3, mice were treated with isotype control, anti-PD-L1 antibody (200 µg i.p. injection every 3 days) and/or PLX3397 (40 mg/kg per os (p.o.) once a day) (n=6 mice/group) until mice met treatment endpoint. (A) Representative images of the Hepa1-6-OPN subcutaneous HCC tumours from each group. (B) Growth of subcutaneous Hepa1-6-OPN tumours. (C) The average tumour weight of each group at the time of sacrifice. (D–F) BALB/C mice were orthotopically implanted H22-OPN tumours with OPN overexpression. (D) Representative images of the H22-OPN orthotopic HCC tumours from each group (n=5 mice/group). (E) Tumour weight at the endpoint of figure part D. (F) Survival curves of figure part D. (G and H) Representative images of H&E staining of lung tissues (G) and the number of lung metastatic foci (F) from each group shown. *P<0.05, **p<0.01 and ***p<0.001, Student’s t-test (or log-rank test in F). Data represent means±SEM. CSF1, colony stimulating factor-1; CSF1R, CSF1 receptor; HCC, hepatocellular carcinoma; OPN, osteopontin.
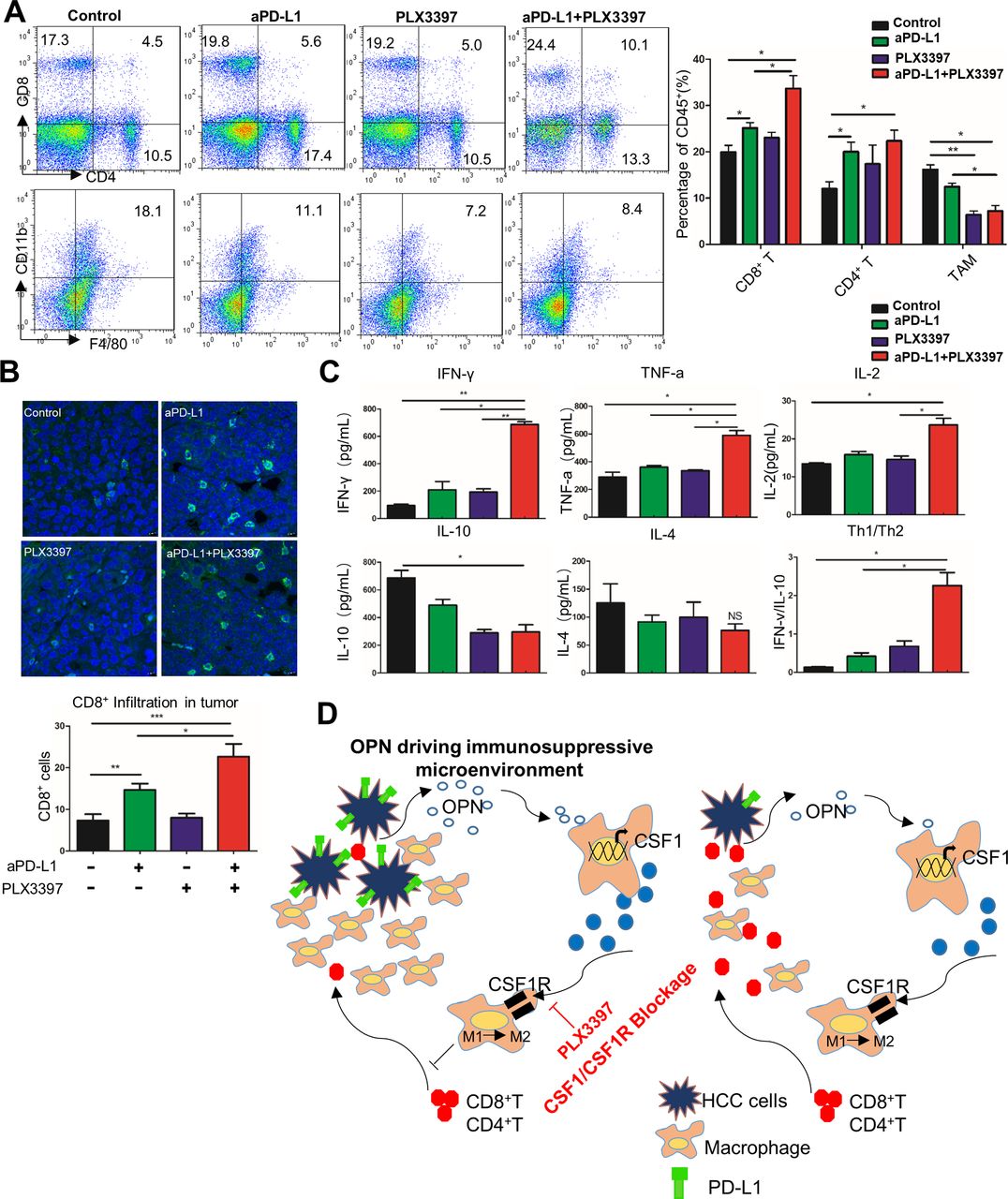
PLX3397 plus PD-L1 antibody reduces TAM infiltration and activates CD8+ T cells in HCC microenvironment
To further understand the antitumour immune response by PLX3397 and anti-PD-L1 combination, we analysed the number of tumour-infiltrating immune cells in Hepa1-6 subcutaneous tumours. Flow cytometric analysis of Hepa1-6 tumours revealed a significant reduction of TAMs in mice treated with the PLX3397 and anti-PD-L1 combination compared with the control (p<0.01, figure 7A and online supplementary figure 8F). Moreover, the percentage of CD8+ T and CD4+ T cells was higher in anti-PD-L1-treated mice than control mice (figure 7A). Importantly, in the combination treatment group, the infiltrating CD8+ T cells were increased as compared the PLX3397 or anti-PD-L1 alone (figure 7A). IHC staining also showed a decrease in F4/80+ TAMs and an increase in CD8+ T and CD4+ T cells in Hepa1-6 tumours after the PLX3397 and anti-PD-L1 combination treatment (see online supplementary figure 8G). Similarly, decreased TAM infiltration and increased intratumoural penetration of cytotoxic CD8+ T and CD4+ T cells were also observed in the H22 orthotopic HCC tumours (figure 7B and online supplementary figure 9A).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
CSF1R inhibitor and anti-PD-L1 combination decreases TAM infiltration and increases intratumoural T cells activation. (A) After study endpoint, the CD4+, CD8+ tumour-infiltrating T lymphocytes (TILs) and F4/80+CD11b+ TAMs in Hepa1-6-OPN subcutaneous tumours from each group were measured by flow cytometry. Left: representative flow cytometry data showing the proportion of CD4+, CD8+ TILs and TAMs in tumour tissues from C57BL/6 mice. Quantification shown on the right. (B) Top: representative images of immunofluorescence confocal microscopy of CD8+ T cells (FITC, green) from H22-OPN orthotopic tumours. Quantification shown on the bottom. (C) The levels of indicators of T cells activation: Th1 cytokines (IFN-γ, TNF-α, IL-2), Th2 cytokines (IL-4 and IL-10) and Th1/Th2 radio (IFN-γ/IL-10) in each group from subcutaneous HCC model were analysed by ELISA. (D) A schematic diagram depicting the microenvironment by CSF1/CSF1R blockade in OPNhigh HCC. OPN facilitates chemotactic migration of monocytes/macrophages and M2-like polarisation of macrophages in the HCC microenvironment. OPN also promotes PD-L1 expression via the activation of the CSF1/CSF1R pathway in macrophages, which enhances the production of immunosuppressive cytokines and drives HCC metastasis through successful evasion of type I INF-dependent or Th1-dependent tumour elimination. Inhibition of the CSF1/CSF1R pathway, for example, PLX3397, reduces macrophage recruitment and M2 phenotype polarisation, which subsequently activates CD8+ T cells and sensitises immune-dependent OPNhigh HCC to anti-PD-L1 immune checkpoint blockade. *P<0.05, **p<0.01 and ***p<0.001, Student’s t-test. Data represent means±SEM. CSF1R, colony stimulating factor-1 receptor; IFN-γ, interferon γ; IL, interleukin; OPN, osteopontin; PD-L1, programmed death ligand 1; TAM, tumour-associated macrophage; TNF-α, tumour necrosis factor α.
To validate the activation of the tumour-infiltrating CD8+T cells after PD-L1 blockade, we evaluated the biomarkers indicative of CD8+ T cells activation in mice plasma at the study endpoint after treatment. Increased levels of IFN-γ, TNFα and IL-2 from activated CD8+ T cells were observed when mice were treated the combination treatment (figure 7C). PLX3397 or anti-PD-L1 monotherapy only weakly activated CD8+ T cells (figure 7C). We previously reported that CSF1 induces a global Th1/Th2-like cytokine shift (an increase in Th2 and decrease in Th1 cytokines) that promotes HCC metastasis.36 To explore the possibility that CSF1/CSF1R blockade could reverse the Th1/Th2 cytokine profiles, we analysed the levels of Th2 cytokine secretion by ELISA. Indeed, the combination of anti-PD-L1 and PLX3397 significantly increased the Th1/Th2 cytokine ratio (IFN-γ/IL-10) compared with each agent alone (figure 7C). Similarly, the PLX3397 and anti-PD-L1 combination also increased the expression of IFN-γ, TNFα and IL-2, as well as Th1/Th2 cytokine balance in H22 orthotopic HCC mice (see online supplementary figure 9B).
On the basis of these findings, we proposed a model shown in figure 7D. OPN facilitates chemotactic migration of monocytes/macrophages and M2-like polarisation of macrophages in the HCC microenvironment. Furthermore, OPN-activated macrophages secrete CSF1 through PI3K/AKT/NF-κB/p65 pathway to drive macrophage into M2-like and promote PD-L1 expression in HCC, which enhances the production of immunosuppressive cytokines in macrophages and drives HCC metastasis through successful evasion of Th1-dependent tumour elimination. Blocking the CSF1/CSF1R pathway reduces macrophage recruitment and M2 phenotype polarisation, which subsequently activates CD8+ T cells and sensitises immune-dependent OPNhigh HCC to anti-PD-L1 immune checkpoint blockade.
Discussion
Gene expression profiling from our previous study identified OPN as one of the leading genes that promotes HCC metastasis.22 Budhu et al 36 found that metastatic HCC exhibited a significant increase in the anti-inflammatory cytokines such as IL-4, IL-5, IL-8 and IL-10 (Th2 cytokines) and a concomitant decrease in proinflammatory cytokines, for example, TNFα, IFN-γ and IL-12b (Th1 cytokines) compared with the normal liver. Such Th1 to Th2 cytokine profile switch accompanied by overexpression of CSF1 is unique to hepatic tissues from metastatic HCC patients and is not related to the degree of viral hepatitis or cirrhosis, or a result of tumour burden.36 Therefore, we propose that a crosstalk between OPN overexpression and CSF1-mediated Th1 to Th2 switch exists and plays a vital role in HCC progression.
Although PD-1/PD-L1 immune checkpoint blockade has demonstrated clinical success,9 only fewer than a quarter of treated patients achieve durable responses, suggesting that the deficiency may be closely associated with the potent pre-existing immune suppressive effects produced by the infiltrating immune cells, such as TAMs.38 39 Expansion of TAMs is a major mechanism used by cancers to escape immune surveillance. Lim et al 40 showed that macrophage-secreted TNF-α increased PD-L1 expression in breast cancer through TNF-α-mediated activation of NF-kB. In this study, we uncovered an OPN-dependent infiltration of TAMs and upregulation of immune checkpoint PD-L1 as a potential mechanism by which HCC escapes immune surveillance. Importantly, our results from the OPN KO model demonstrated that OPN facilitates M2-like polarisation of macrophages and promotes the PD-L1 expression in HCC, which resulted in selection of tumours producing tumour-promoting factors that blocked Th1 immunity and initial type I IFN-dependent innate immune activation against tumours. Moreover, analysis of a clinical HCC data set also supported a positive association between OPN, PD-L1 and TAM infiltration as well as related gene expression by the M2-like macrophage activation and PD-L1 upregulation in OPNhigh HCC cocultured with macrophages in vitro. CSF1 secretion by TAMs likely plays an important role given that CSF1 was one of the highly increased cytokines.
The next frontier of immunotherapy is to discover ways to enhance the efficacy of checkpoint inhibitors by combination approaches, especially in cancers in which limited clinical effects are observed. In particular, recent studies indicated that the combination of anti-PD-1/PD-L1 and cxyclooxygenase-2 inhibitor, epigenetic drugs, radiotherapy or ablation therapy synergistically enhances antitumour immunity,41–44 suggesting that antitumour agents that can induce tumour-specific T cells activation could deliver promising results in combination with PD-1/PD-L1 immunotherapy.
In that regard, we propose that CSF1/CSF1R blockade has the potential to modulate the immunological features of HCC and enhance the efficacy of anti-PD-L1 blockade. Finally, our data showed that CSF1R inhibitors significantly reduced tumour growth when combined with anti-PD-L1 blockade in mice. Furthermore, we showed that combination treatment inhibited the recruitment of inflammatory monocytes, and infiltration and M2-polarisation of TAMs, resulting in reversal of the immunosuppression status of HCC microenvironment and activation of an antitumour CD8+ T cells response. CSF1/CSF1R-targeting agents have been investigated in multiple tumour types.45–47 Recently, Pyonteck et al 48 reported that blockade of CSF1R signalling, using a small-molecule inhibitor BLZ945, significantly inhibited murine glioma tumour growth by reprogramming TAM response. In addition, Neubert et al 49 demonstrated that anti-CSF1R–mediated macrophage elimination may be beneficial in melanomas refractory to immune checkpoint blockade. Those findings are in line with the current study.
A key issue with DEN/CCL4 mice is the fact that OPN KO mice are whole body KO mice and caused the delayed tumour growth; we really cannot exclude that tumour intrinsic effect are involved in the OPN-CSF1 axis. Considering OPN knockdown can induce a significant decrease in HCC proliferation and the delayed tumour growth may have an effect on the reduced macrophage infiltration and PD-L1 expression, it is difficult to dissect the effects driven by OPN directly or by OPN in tumour growth in vivo. However, our results (figure 3A–F and online supplementary figure 5A–D) clearly demonstrate that OPN induces the macrophage migration and polarisation and promotes PD-L1 expression in HCC through the effect on macrophages. Then, it is worth noting that OPN KO mice have lower PD-L1 expression within tumours, but have high IFN-γ expression, despite of the fact that IFN-γ has been reported to be the main factor of PD-L1 upregulation. It is likely that OPN KO may affect IFN-γ receptor (IFNGR)/STAT324 and thereby affect IFN-γ-mediated PD-L1 upregulation. Although our experiment detected differences in PD-L1 expression in HCC cells exclusively between OPN KO and WT mice, PD-L1/PD-1 ligation between haematopoietic cells and tumour may also contribute to immunosuppression. Because the combination is more effective than single treatment, CSF-1 and PD-L1 may work independently to suppress the immune cell. Considering the PD-L1 also expressed in other immune cells that do not respond to anti-CSF1, their combination can exert better therapeutic outcomes.
In conclusion, the current results identifying the OPN/CSF1/CSF1R axis as a critical mediator in the immunosuppressive nature of the HCC microenvironment suggested that targeting this pathway may prevent TAM trafficking and improve the efficacy of immune checkpoint blockade in HCC.
References
Footnotes
YZ, JY, DX, X-MG and ZZ contributed equally.
Contributors YZ, JY, DX, XM-G and ZZ designed and performed the experiments, analysed data and wrote the manuscript; JH, LL and HL-J revised the manuscript; JH-L, YY-S, QL and YZ performed experiments and analysed data; CW-L and SO-L participated in the discussion and data interpretation; YZ and YZ analysed data; LX-Q, QZ-D and MC-H designed and supervised the entire project, designed the experiments, analysed data and wrote the manuscript.
Funding This work was supported by the following: the National Key Research and Development Program of China (2017YFC1308604); the National Key Basic Research Program of China (2014CB542101 and 2013CB910500); National Natural Science Foundation of China (81772563, 81672820, 81372647 and 81672365); and China National Key Projects for Infectious Disease (2012ZX10002-012 and 2017ZX10203207).
Competing interests None declared.
Provenance and peer review Not commissioned; externally peer reviewed.
Patient consent for publication Not required.