Article Text

Abstract
Background: Previous studies have reported that soluble (s) CD86 is involved in the initiation of the immune response. A study was undertaken to investigate the concentrations of sCD86 in serum samples from patients with bronchial asthma and to determine the cell origin of sCD86.
Methods: Serum sCD86 concentrations were measured in 52 asthmatic subjects and 25 non-atopic normal volunteers using an enzyme linked immunosorbent assay, and the relationship of serum sCD86 concentrations to asthma severity and to total and differential white cell counts was analysed. Each type of white blood cell was purified and cultured in vitro to determine the cell origin of serum sCD86.
Results: Serum samples from patients with an acute asthma exacerbation had much higher levels of sCD86 (585.4 (20.5) IU/ml) than those from stable asthmatics (479.6 (15.7) IU/ml, p<0.001) and healthy individuals (435.1 (13.8) IU/ml, p<0.001), and there was no difference between the latter two groups (p = 0.079). In asthmatic subjects the serum sCD86 level was inversely correlated with airway responsiveness, forced expiratory volume in 1 second, and with arterial carbon dioxide tension. In addition, the serum sCD86 level was positively correlated with numbers of lymphocytes, eosinophils, monocytes, but not neutrophils. The in vitro experiments indicated that sCD86 was produced by monocytes.
Conclusions: The serum sCD86 protein level was significantly increased in asthmatic subjects during an exacerbation and correlated with the severity of asthma. sCD86 is most probably derived from monocytes in the peripheral blood.
- soluble CD86
- asthma
- pathogenesis
Statistics from Altmetric.com
Naïve T lymphocytes require two distinct signals from antigen-presenting cells (APC) to be functionally activated.1,2 The first signal, which confers specificity, is provided by the interaction of the T cell receptor with major histocompatibility complex II complexes on APCs. A second co-stimulatory signal can be provided by APC borne ligands for the CD28 and cytotoxic lymphocyte associated antigen-4 (CTLA-4) receptors on T cells. CD28 is constitutively expressed by T cells and interacts with the B7 molecules B7-1 (CD80) and B7-2 (CD86).3,4 CD80 and CD86 are capable of forming homodimers, allowing for interactions with homodimers of either CD28 or CTLA-4. The interaction of CD28 with its ligands is weaker than the interaction with CTLA-4. Human CD80 binds to human CTLA-4 and CD28 with Kd values of 0.42 and 4 μM, respectively, for the monomeric interactions.5 The CD86:CTLA-4 interaction is of an affinity similar to CD80:CD28 and the CD28:CD86 interaction is of even lower affinity.6 CD80 and CD86 are type 1 membrane glycoproteins belonging to the immunoglobulin (Ig) supergene family. Despite having the same ligands, CD80 and CD86 appear to be involved in different mechanisms; CD80 can be more potent than CD86 in inducing an antitumoral response while CD86 preferentially induces the production of a helper (Th) 2 response.7,8 Due to its constitutive expression on human APCs, it has been suggested that CD86 may be involved in the initiation of the immune response.
Allergic asthma is a disease characterised by airway hyperresponsiveness, pulmonary inflammation, and raised serum IgE levels. Production of Th2 cytokines such as interleukin (IL)-4, IL-5 and IL-13 in allergic asthma is at least partly responsible for eliciting the cardinal pathogenic changes of the asthmatic phenotype.9 The factors in asthma that govern the production of Th2 cytokines over Th1 cytokines, such as interferon-γ, are slowly being revealed. One likely factor is the cytokine profile of T cells which is influenced by APCs, possibly through co-stimulatory signals. Such signals may be provided by ligation of CD28 or CTLA-4 with CD80 or CD86.10 Jeannin and coworkers have reported that soluble (s) CD86 results from an alternatively spliced transcript characterised by the deletion of the transmembrane domain; and that sCD86 provides a co-stimulatory signal to memory human T cells.11 Circulating sCD86 has been reported to be detected in the plasma of normal individuals and patients with leukaemia.12 Reverse transcriptase polymerase chain reaction analysis has shown that the transcript for sCD86 is expressed in normal monocytes, dendritic cells, as well as some leukaemic cells, but not in normal T, B and NK cells.12 Monocytes in peripheral blood might therefore be able to produce sCD86. We speculated that a reasonable starting point in the search for sCD86 expression would be in patients with asthma. To that end, we determined the concentrations of sCD86 in serum samples from patients with asthma and the correlations between sCD86 levels and clinical indices as well as the cell origin of sCD86.
METHODS
Subjects
The study protocol was approved by our institutional review board for human studies and informed consent was obtained from all subjects. Sixty eight asthmatic subjects were recruited consecutively from regular attenders in the pulmonary outpatient or emergency department at First Affiliated Hospital, Guangxi Medical University, PR China in the year from September 2001 to August 2002. The diagnosis and classification of the severity of asthma were based on the NHLBI/WHO Workshop on the Global Strategy for Asthma (GINA guidelines).13 The minimum criteria for the diagnosis of an asthma exacerbation included intense subjective breathlessness, audible wheezing on auscultation, and a morning peak expiratory flow <70% of the predicted value in the previous 3 months. Atopy was defined as a positive prick test to one or more common aeroallergens. Thirteen asthmatics were excluded from the study because they had been treated with oral and/or intravenous corticosteroids in the preceding 4 weeks, and three patients refused to participate in the study. Finally, 28 patients were recruited during a moderate to severe exacerbation either on their scheduled visit to the pulmonary outpatient department or during an emergency visit, and 24 patients with stable asthma were recruited on their scheduled visit if the symptoms and peak expiratory flow were stable with no change in treatment for at least 1 month. None of the studied patients with asthma smoked, had complications of other lung diseases, or had a history suggesting intolerance to non-steroidal anti-inflammatory drugs. Twenty five non-smoking healthy volunteers were studied as controls. They had no history of atopic factors or allergic diseases, and no evidence of any lung diseases. Table 1 summarises the clinical characteristics and forced expiratory volume in 1 second (FEV1, expressed as a percentage of the predicted value) of the patients on recruitment to the study.
Characteristics of patients with asthma and control subjects
Assessment of airway responsiveness
Baseline measurements of FEV1 were followed by methacholine challenge. Methacholine inhalation tests were carried out by the method described previously14 until a provocative concentration causing a 20% decrease in FEV1 (PC20-MCh) was obtained.
Blood samples
Arterial blood samples were obtained from each subject for blood gas analysis to determine the oxygen and carbon dioxide tensions (Pao2, Paco2). Venous blood samples were collected either in ethylenediamine tetraacetic acid treated tubes for total and differential blood cell counts or untreated tubes to obtain serum for determining sCD86 concentrations, and the serum samples were stored at −70°C before determination. Total cell counts were performed with a haemocytometer and different cell counts were made from blood smears stained with Diff-Quik Staining (Sigma). Cells were classified as lymphocytes, neutrophils, eosinophils, monocytes, and basophils as standard morphological criteria and absolute numbers of each cell type were calculated.
Cell isolation and culture
Heparinised venous blood was drawn from six healthy volunteers and six asthmatics during an acute exacerbation. Granulocytes and peripheral blood mononuclear cells (PBMCs) were isolated by centrifugation on Ficoll-Paque (Pharmacia, Uppsala, Sweden). The neutrophil layer was collected and the remaining erythrocytes removed by hypotonic lysis. Eosinophils were purified by negative immunomagnetic selection using a magnetically activated cell separator system (Miltenyi Biotec, Auburn, California, USA) as previously described.15 By negative selection, highly purified eosinophils, depleted of neutrophils (CD16+ cells) and any contaminating mononuclear cells (CD3+/CD4+/CD19+ cells), were routinely obtained. Untouched T cells, B cells, and monocytes from PBMCs were isolated by negative immunomagnetic selection using a Pan T cell isolation kit II, B cell isolation kit II, and monocyte isolation kit II, respectively (all purchased from Miltenyi Biotec), and all magnetic separation procedures were carried out according to the protocols provided by the manufacturer. The purities of T cells, B cells, neutrophils, eosinophils, or monocytes were all >95%. One million T cells, B cells, neutrophils, eosinophils, or monocytes alone were cultured in a volume of 1 ml complete medium (RPMI 1640 supplemented with 5% fetal calf serum and antibiotics reagent). After 24 hours the cell-free supernatants were collected and stored at −70°C for determining sCD86 concentrations.
Detection of sCD86
The serum and supernatant samples previously stored at −70°C were thawed and approximately 300 μl of each sample were centrifuged for 10 minutes at 16 000×g to remove any cell debris. The concentrations of sCD86 in serum samples and supernatants of cultured cells were measured by a sandwich enzyme-linked immunosorbent assay kit according to the manufacturer’s protocol (Diaclone, Besançon, France). The minimum detectable dose of sCD86 was 0.6 U/ml. All samples were assayed in duplicate.
Statistical analysis
Data were presented as mean (SE). Statistical analysis was done by one-way analysis of variance (ANOVA) for data conforming to a normal distribution, and by Friedman’s test for those data with a non-parametric distribution (confirmed by the Shapiro-Will W test). The Student’s t test was used for comparison of two groups, and the Pearson or Spearman rank correlation was used to analyse correlations. A p value of <0.05 was considered statistically significant.
RESULTS
Clinical characteristics
Background data for the 25 normal control subjects, 28 patients with acute asthma exacerbations, and 24 with stable asthma enrolled in the study are shown in table 1. The three groups did not differ significantly in age or sex (both p>0.05). There were no significant differences in atopic status or duration of asthma between patients with acute asthma exacerbations and those with stable asthma (both p>0.05). %FEV1 in asthmatic subjects during an acute exacerbation decreased significantly compared with those with stable asthma and control subjects (p<0.001).
Serum sCD86 levels in patients with asthma
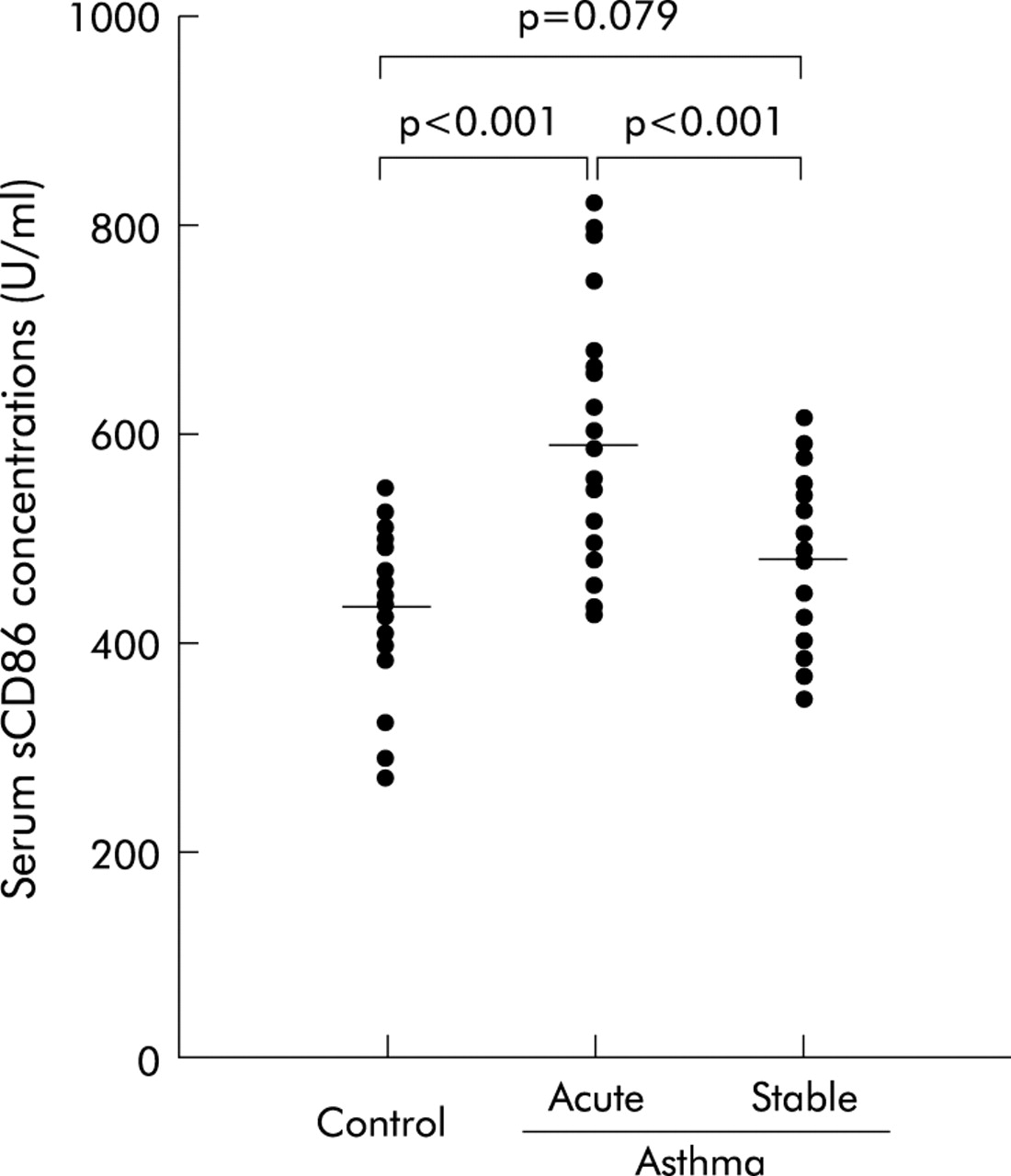
Figure 1 shows combined data of serum sCD86 concentrations from normal healthy volunteers, patients with acute asthma exacerbations, and those with stable asthma. sCD86 was detectable in all subjects studied. There was no obvious relationship between levels of sCD86 and the sex or age of the subjects. Serum samples from patients with acute asthma exacerbations had much higher levels of sCD86 (585.4 (20.5) U/ml (95% confidence interval (CI) 543.4 to 627.4)) than serum from stable asthmatics (479.6 (15.7) U/ml (95% CI 447.1 to 512.1) p<0.001) and healthy individuals (435.1 (13.8) U/ml (95% CI 406.6 to 463.6) p<0.001), and there was no difference in serum sCD86 concentration between stable asthmatics and healthy individuals (p = 0.079). In addition, the serum sCD86 concentration in 36 patients with atopic asthma (531.8 (17.7) U/ml (95% CI 495.8 to 567.8) did not differ from that in 16 patients with non-atopic asthma (547.2 (28.8) U/ml (95% CI 485.9 to 608.5), t = 0.469, p = 0.641).

Comparison of serum concentrations of sCD86 in normal control subjects and asthmatic patients.
Correlation between serum sCD86 levels and various clinical parameters
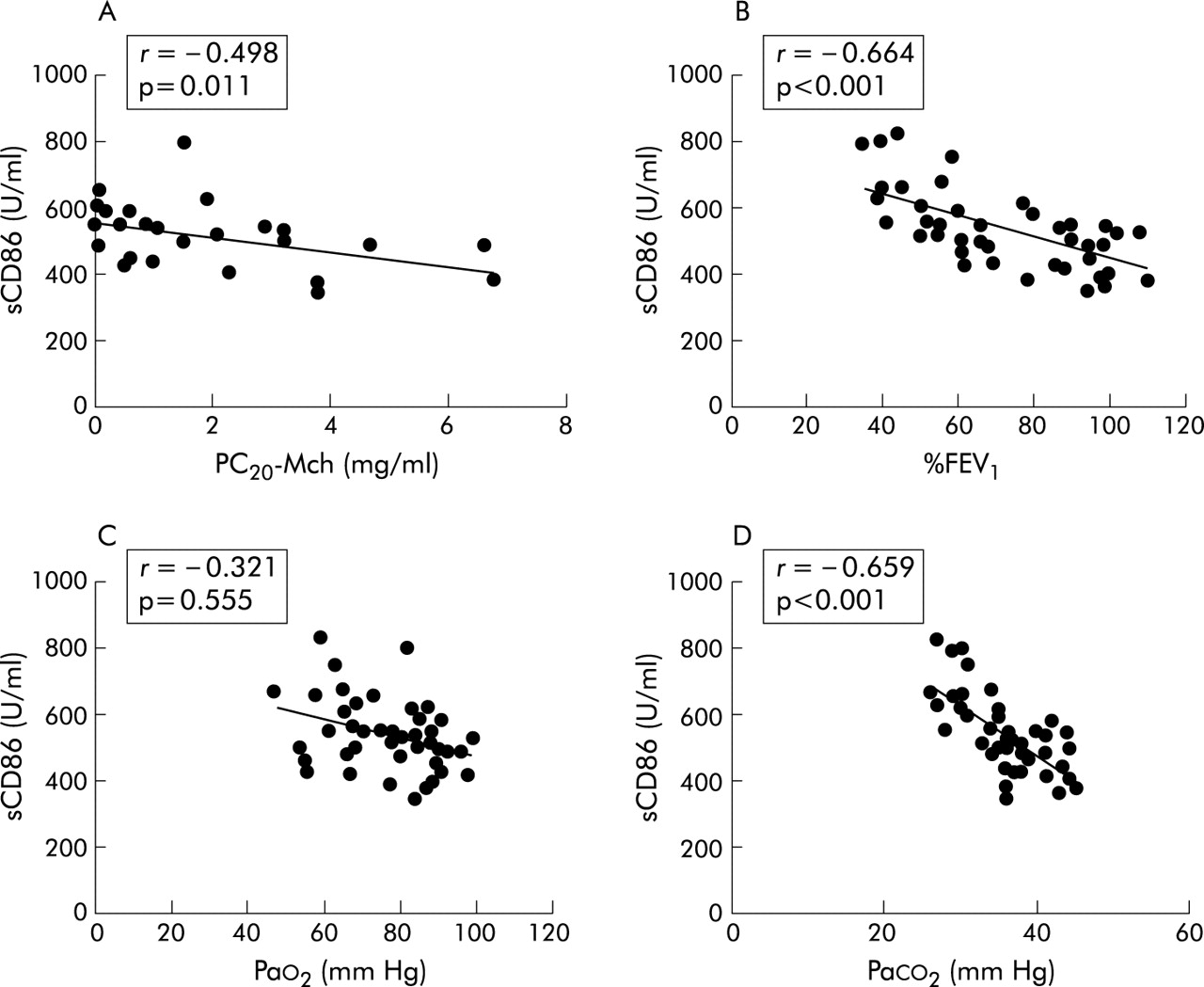
Of 52 asthmatic patients, %FEV1 was more than 60% in 38 and airway responsiveness was measured in 25 of these (20 atopic and five non-atopic asthmatics). The geometric mean (SE) PC20MCh was 0.89 (1.07) mg/ml. Details of %FEV1, Pao2 and Paco2 in the study subjects are shown in table 1. The serum sCD86 level was inversely correlated with PC20-MCh (r = −0.498, p = 0.011), % FEV1 (r = −0.664, p<0.001), and with paco2 (r = −0.659, p<0.001, fig 2), but did not correlate with Pao2 (r = −0.321, p = 0.555). Total and differential blood cell counts in healthy controls and asthmatic subjects are shown in table 2. In 52 patients with asthma the serum sCD86 level was positively correlated with numbers of lymphocytes (r = 0.628, p<0.001), eosinophils (r = 0.659, p<0.001), or monocytes (r = 0.729, p<0.001) but not neutrophils (r = −0.135, p = 0.339; fig 3).
Mean (SE) white blood cell counts and differential counts in asthmatic and control subjects

Correlation between concentrations of serum sCD86 and (A) PC20-MCh (n = 25), (B) %FEV1 (n = 52), (C) Pao2 (n = 52), and (D) Paco2 (n = 52) in patients with asthma.

Correlation between concentrations of serum sCD86 and numbers of (A) lymphocytes, (B) neutrophils, (C) eosinophils, and (D) monocytes in peripheral blood of 52 patients with asthma.
Cell origin of sCD86
To reveal the cell origin of sCD86, untouched T cells, B cells, neutrophils, eosinophils, and monocytes from peripheral blood were purified and cultured to determine the presence of sCD86 in the supernatants. No sCD86 could be detected in the supernatants of cultured T cells, B cells, neutrophils, or eosinophils from either asthmatic subjects or healthy volunteers. The most important finding was that monocytes were the only cell type which released sCD86 into the culture supernatants, and that monocytes from asthmatic subjects produced more sCD86 than those from healthy volunteers (fig 4).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
sCD86 production by non-activated monocytes in vitro. One million monocytes from six normal control subjects and six asthmatic subjects with an exacerbation were purified and cultured in a volume of 1 ml for 24 hours and the supernatants were collected for determination of sCD86 concentrations.
DISCUSSION
We have shown that sCD86 is detected in the serum of healthy controls and both atopic and non-atopic asthmatic subjects, and that sCD86 levels are raised in the serum of patients with acute asthma exacerbations. The concentration of serum sCD86 in asthmatic patients was inversely correlated with PC20-MCh, %FEV1, and Paco2. These findings suggest a potentially important role for sCD86 in asthma. To our knowledge, this study is the first to suggest the possible clinical significance of sCD86 in human asthma.
Co-stimulation through the B7/CD28 pathway plays a critical role in the establishment of antigen driven immune responses.16 Hofer and coworkers17 found that atopic patients with asthma who were exposed to allergens had significantly higher levels of CD86 expression on B cells than atopic asthmatic subjects not exposed to allergen in vivo or non-atopic controls. In contrast, there were no differences in CD80 expression among the three study groups. When PBMCs from asthmatic patients or normal control subjects were stimulated with IL-4 or IL-13, the expression of CD86 (but not CD80) was significantly increased on B cells.17 It has been shown that blocking CD86 but not CD80 co-stimulation is effective in inhibiting allergic responses and that these effects are associated with reduced Th2 cytokine production.18,19 Tsuyuki and coworkers20 have also reported that airway allergen challenge upregulates the expression of CD86 on B cells from the lung and airway administration of anti-CD86 monoclonal antibody inhibits local IgE production in a mouse model of asthma. In addition, blockade of co-stimulation with CTLA4-Ig (a fusion protein known to prevent co-stimulation by blocking CD28/B7 interactions) inhibits airway hyperresponsiveness, inflammatory cell infiltration, and allergen-specific responsiveness of thoracic T cells in a murine model of asthma.21 CD28-deficient mice did not manifest airway inflammation in response to allergen challenge, despite evidence of systemic sensitisation, and this was due to a failure in T cell recruitment and defective Th2 cell development.22 Prevention of CTLA-4 engagement by blockade of CD86 in CD28-deficient mice restored lymphocyte recruitment to the airway but did not result in tissue eosinophilia, Ig isotype switching, or Th2 cytokine secretion.22 All the abovementioned data support the hypothesis that CD86 is an important co-stimulatory molecule in the pathogenesis of allergic asthma.
Numerous studies have shown that soluble forms of B7 molecules have potent functional activity and that, depending on the experimental system used, can either inhibit or enhance immune responses.11,23,24 sCD86 might play an important role in immune regulation by binding with the CD28 molecules, thus interfering with the binding of CD28 and/or CTLA-4. Recombinant sCD86 at high concentrations in vitro could inhibit mixed leucocyte reactions and cellular proliferation,25 probably by interference with CD28 signalling. sCD86 might also interfere with CD28-B7 interactions and block negative signals transmitted via surface bound CD86. It has been reported that sCD86 results from an alternatively spliced transcript (CD86deltaTM) characterised by deletion of the transmembrane domain.11 Recombinant CD86deltaTM binds to CD28 and CTLA-4 and induces the activation of T cells after stimulation with anti-CD3 monoclonal antibody. CD86deltaTM also induces interferon-γ production by virus specific CD8+ memory human T cells stimulated with the Flu M1 peptide.11 sCD86 molecule therefore appears to be a functional co-stimulatory molecule playing a potentially important role in immune surveillance. The high levels of sCD86 in serum samples from asthmatic subjects with acute exacerbations may play a protective role via interference with the CD28-B7 interaction or, alternatively, contribute to disease manifestations by interference with the CD28-B7 interaction.
Membrane CD86 is expressed predominantly on monocytes, dendritic cells, lymphocytes, eosinophils, and neutrophils.11,26–29 It has been found that the transcript for sCD86 is expressed in normal monocytes and dendritic cells.12 Our results showed that the serum sCD86 level was positively correlated with numbers of lymphocytes, eosinophils, and monocytes, but not neutrophils. These results suggest that sCD86 detected in asthmatic serum might derive from lymphocytes, eosinophils, and monocytes. However, our in vitro experimental results showed that cultured T cells, B cells, neutrophils, or eosinophils from either asthmatics or healthy volunteers did not produce detectable sCD86. In contrast, without any stimulation, monocytes were capable of releasing sCD86 into the culture supernatants. We also noted that monocytes from asthmatics produced more sCD86 than those from healthy volunteers. We therefore infer that serum sCD86 is derived from monocytes in peripheral blood and that monocytes are further responsible for the increase in serum sCD86 in asthmatic patients with acute exacerbations. On the other hand, we cannot rule out the possibility that other types of leucocytes such as lymphocytes and eosinophils are also able to produce sCD86 in vivo under asthmatic conditions.
In summary, our results show that the serum sCD86 protein level is significantly increased in asthmatic patients during an exacerbation and that this level correlates with the severity of asthma. Our results also suggest that sCD86 is probably derived from monocytes in the peripheral blood. Further elucidation of the roles and functions of sCD86 in bronchial asthma will require longitudinal studies and detailed molecular research, comparing the immunopathological characteristics of asthma in diverse clinical settings using samples of induced sputum, bronchoalveolar lavage fluid, or bronchial biopsy specimens.
Acknowledgments
The authors thank Miss Jian Li for her skilful technical assistance, and Feng Gao for helpful discussions and critical review of the manuscript.
REFERENCES
Footnotes
-
This study was supported in part by research grant no 30060079 from the National Natural Science Foundation of China, in part by research grant no 200260 from the Ministry of Education, People’s Republic of China, and in part by research grant no 200203 from the Bureau of Health, Guangxi Zhuang Autonomous Zone, People’s Republic of China.